«Границы моего языка определяют границы моего мира». Витгенштейн.
Наиболее впечатляющие достижения человека связаны с теми видами поведения, коюрые обеспечивают его коммуникационную деятельность. В четвёртой части этой книги рассматривается структура коммуникации у приматов исходя из двухпроцессного механизма функционирования мозга, неврологической организации восприятия и чувств, а также регулирующего воздействия нервной системы на поведение. В главе XVI указывалось, что коммуникация предполагает иерархическую взаимосвязь сенсорных процессов органов чувств и ствола мозга с моторными механизмами. В последующих главах эта взаимосвязь будет рассматриваться детально. Можно выделить два вида коммуникационных актов исходя из того, зависит ли смысл действия от ситуации, в которой оно происходит. Коммуникационные акты, не зависящие от ситуации, мы будем называть «знаками», и их неврологическая организация рассматривается в данной главе. Коммуникационные акты, зависящие от ситуации, мы будем обозначать термином «символы», механизмы мозга, ответственные за их образование, детально описываются в Во всех этих главах мы будем исходить из того, в какой мере эти процессы зависят от ситуапии. По существу, предполагается, что знаки не зависят от контекста, они являются результатом воздействия ассоциативной коры (см. Pис. Значение символа, напротив, определяется контекстом. Таким образом, широко распространённый символ мира в других контекстах может быть символом победы, приветствия и так далее. Существует определённое сходство между символами и другими зависящими от контекста типами поведения, например теми, которые определяют взаимоотношения людей, так как Пользуясь в этих главах терминами знак и символ, мы исходим из обычного их употребления в философии (например, Ауег, 1946; Morris, 1946; Langer, 1951) Что такое «знак»Когда я начал проводить опыты на обезьянах, я обычно применял в качестве подкрепления правильного выбора, ведущего к решению проблемы, очищенный арахис. При подготовке к серии опытов приходилось тратить много времени на очистку орехов. Внезапно я понял, что нет оснований считать, что обезьяны не смогут сами чистить орехи, и действительно оказалось, что они справляются с этим, не задерживая процедуры эксперимента. Меня интересовала выработка дифференцировок, Эти ранние эксперименты убедили меня в том, что выработка дифференцировки представляет собой изучение способности организма раскрывать смысл знаков. Этой формулировке, однако, не хватало той строгости, которую принято связывать с результатами эксперимента. Ведь «знак», который чётко указывает на Эта неясность была устранена сразу благодаря удивительным результатам, полученным Робертом и Беатрис Гарднер (1969), работавшими с молодой самкой шимпанзе по кличке Уошо. Поскольку попытки научить шимпанзе речи не удались, они решили попытаться научить её общаться при помощи рук, а не голосовых связок. Они остановили выбор на американской системе знаков, применяемой глухонемыми, считая эту систему идеальным средством, поскольку она легко поддаётся изучению благодаря своей наглядности, ведь в этой системе при помощи кистей рук и пальцев создаётся некоторое подобие предмета или действия, обозначаемого этим знаком. Уошо, которой в то время, когда писалась эта книга, было около трёх с половиной лет, заучила примерно 150 таких знаков. Гарднеры и Уошо легко общались друг с другом, используя этот метод. Фактически Уошо даже изобрела несколько новых знаков, которые обычно употребляются людьми, обучающимися этой системе. Мы ещё несколько раз вернёмся в следующих трёх главах к этому имеющему большое значение эксперименту с Уошо. Здесь же нам нужно обсудить только один основной факт, то, что приматы могут создавать знаки и сообщать при помощи знаков, независимых от контекста, об устойчивых признаках ситуации, которые они могут таким образом дифференцировать и узнавать. А о механизмах мозга, участвующих в выработке дифференцировок и узнавания, нам известно очень много. Выработка дифференцировок и узнаваниеНекоторые аспекты распознавания образов обсуждались в главах VI, VII и VIII. Участие механизмов памяти описывалось как образование нейронного экрана, включающего ряд фильтров и голографических микроструктур, посредством которых и на которые проецируется вводимая информация для создания Образа. В данной главе мы займёмся детальнее некоторыми из факторов, влияющих на этот зкран во время обучения и опознавания. В главах VI, VII и VIII процесс формирования Образа рассматривался как сравнительно пассивный механизм, который приводится в действие поступающей информацией. Однако ранее мы останавливались и на некоторых активных аспектах этого процесса. Например, в
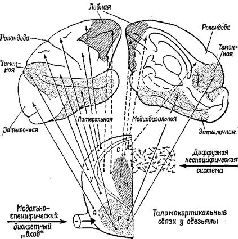
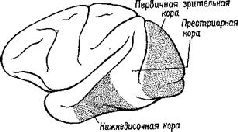
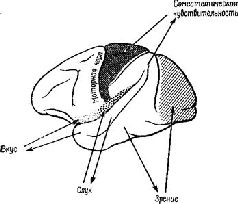
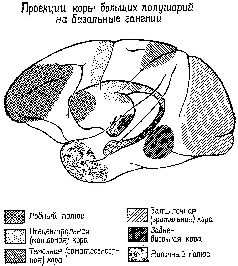
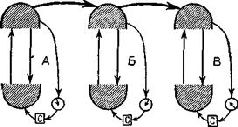
Образ в нашем понимании является — целостным и топографическим. Однако, когда мы в действительности смотрим на окружающий мир или слушаем его, мы избираем тот или иной аспект, мы Следовательно, заучивание и опознавание знака требуют В этой главе мы утверждаем, что знаки создаются тогда, когда наши действия влияют на воспринимаемые Образы. Знак — это действие, представляющее воспринимаемый Образ. Выработка дифференцировок, распознавание образов, избирательное внимание — всё это требует участия нервных механизмов выбора, а этот выбор и запускает в ход действия, которые в свою очередь изменяют созданный Образ. Это положение является итогом серии экспериментов на обезьянах, которые опрокинули представления о том, каким образом функционируют так называемые ассоциативные зоны мозга приматов. Т. Мейнерт ( В частности, множество экспериментов, проведённых на обезьянах, показало, что выработка дифференцировок и распознавание образов возможны и после удаления больших участков коры, окружающей первичные проекционные зоны (Pribram, Spinelli and Reitz, 1969, Для исследователей, стоящих на позициях ассоцианизма, эти: результаты кажутся ещё одним из тех парадоксов, которые так стимулировали исследования в области нервных механизмов поведения на протяжении последних полутора столетий. Каким образом Эта загадка была частично разрешена, когда было установлено, что результаты разрушения мозга неоднозначны. Локализация функций различных сенсорных модальностей была обнаружена и внутри так называемой ассоциативной коры. Один участок в теменной доле мозга оказывает исключительное влияние на различение кожно-кинестетических сигналов; другой участок в передней части височной доли имеет отношение только к вкусовым ощущениям. Средневисочные участки коры специализированы исключительно на слухе, а участок в нижней части височной доли имеет отношение только к зрению (см. Pис. Таким образом, проблема сводится к определению функций этих специфических сенсорных зон. Экспериментальные исследования для разрешения зтой проблемы проводились в основном на зрительных процессах и поэтому касались коры нижней части височной области. Однако Прежде всего возник вопрос, вызывают ли повреждения нижневисочной коры мозга обезьян нарушение всех форм зрительных навыков. Скоро удалось установить, что такое повреждение не нарушало слежения за зрительным объектом, например за летящим комаром. Позднее это было подтверждено посредством фотографирования отражения наблюдаемых объектов от роговицы (Bagshaw, Mackworth and Pribram, 1970). Нарушения возникали лишь в том случае, когда перед обезьянами с повреждениями мозга стояла задача выбора или дифференцировки. Такие нарушения были обнаружены при различных формах зрительного выбора независимо от того относился ли он к цвету, форме, контуру или яркости (Mishkin and Pribram, 1954). Единственным общим в этих задачах было то, что все они были зрительными и во всех предусматривался выбор, а также то, что массивность нарушения поведения была пропорциональна трудности, которую испытывали при обучении нормальные обезьяны. Эти результаты сразу же заставили исследователей предположить, что затруднения, наблюдаемые у обезьян с повреждениями мозга, выражаются скорее в их неспособности заучить (и запомнить) задачу, чем в неспособности воспринять основные ключевые признаки, которые ведут к её разрешению. Для проверки этой гипотезы было проведено исследование того, как обезьяны с удалением нижневисочной коры Следует, однако, обратить внимание на то, что здесь сказано «в общем». Расхождение между влияниями, оказываемыми этими двумя типами повреждений на восприятие и научение, не было ни полным, ни вполне определённым. Результатом повреждения первичной коры было некоторое замедление процесса научения, что можно было ожидать, так как обезьяны приспосабливались к сниженной остроте зрения и большим скотомам в зрительном поле. Более загадочным было то, что у обезьян с повреждениями нижневисочной коры наблюдалось некоторое снижение остроты зрения, например в отношении размера (Mishkin, Hall, 1955) и критической частоты мельканий (Mishkin and Weiskrantz, 1959). Поскольку острота зрения проверялась методом выработки дифференцировок, эта загадка легко объяснялась тем, что был нарушен механизм выбора, Как мозг контролирует поступающую информацию
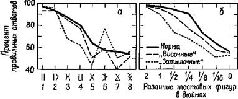
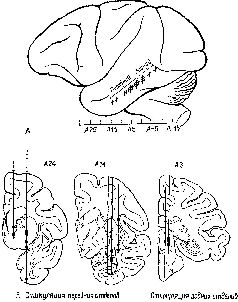
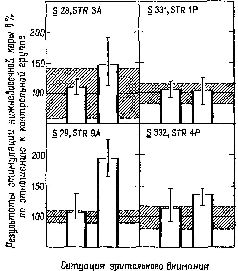
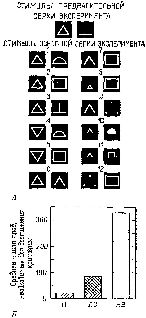
Мы были озадачены тем, что вначале не могли представить себе механизм, который частично — но не полностью — разделял бы восприятие и научение. В классическом понимании функций коры вводимая информация воспринимается посредством первичной коры; к тому, что уже заучено, добавляется затем сумма других видов информации и ассоциаций, хранимых в ассоциативной коре. У. Пенфилд (1969) Как указывалось во введении к этой главе, можно полностью разрешить этот парадокс, если серьёзно рассмотреть гипотезу о том, что влияние нижневисочной коры на зрительные процессы обусловлено не столько тем, что она получает зрительную информацию от первичной коры, сколько тем, что она воздействует через кортикофугальные связи на процессы переработки зрительной информации, происходящие в подкорковых структурах. Какие же факты делают вероятным существование такого коркового контроля подкорковых механизмов зрения? И как можно представить себе его функционирование на основании уже известных нейропсихологических данных? Вспомним, что подрезка нижневисочной коры, прерывающая связи между этой зоной и подкорковыми уровнями, вызывает такое же нарушение функции, как и удаление самой этой зоны. Куда же ведут эти связи? Путь нервных волокон, идущих от нижневисочной коры, был прослежен при помощи анатомических (Whitlock and Nauta, 1954) и физиологических (Reitz and Pribram, 1969, Влияние электрической стимуляции нижневисочной коры может быть зарегистрировано на различных уровнях проводящих путей. Изменения электрической активности, вызванные вспышками света, можно получить и путём раздражения височной коры. Эти изменения наблюдались в первичной коре, в латеральном коленчатом теле, таламусе и даже в зрительном нерве. Связи, идущие от базальных ганглиев и двухолмия к зонам таламуса и сетчатки, не были полностью исследованы, но было сделано достаточно, чтобы установить существование таких связей, хотя в отношении контроля мозга над функциями сетчатки (и, следовательно, зрительного нерва) пока имеются только электрофизиологические данные (Spinelli and Pribram, 1966, 1967, Более полно были прослежены проводящие пути от височной коры к периферии в слуховой системе. И снова двухолмие (в данном случае слуховое) представляет собой важное место переключений; от него проводящий путь нисходит к кохлеарному ядру (либо непосредственно, либо через верхнюю оливу, либо обоими путями; Nobel, Dewson, 1966). Таким образом, снимается вопрос о том, существуют ли связи, посредством которых так называемая ассоциативная кора может оказывать влияние на поступающую информацию. То, что этот контроль тесно связан с двигательными механизмами, согласуется с данными нейропсихологии, говорящими о том, что височная кора играет некоторую роль в тех случаях, когда следует произвести активный выбор. Но каким образом этот выбор влияет на классификацию поступающей информации, на создание знаков и придание им значения? ВниманиеОдной из самых поразительных особенностей Образов является их богатство. Кк правило, всё это богатство нельзя обнаружить сразу, его нужно выявлять постепенно, признак за признаком. Выделяется то цвет, то форма, то структура воспринимаемых объектов. Однако это ограничение процесса внимания не ослабляет непосредственность и калейдоскопическую природу Образа — пожалуй, и то и другое возрастает при надлежащем внимании. Каким же образом? Ответ нужно, конечно, искать в непрерывном взаимодействии между процессом формирования Образа и вниманием. Для осуществления этого имеется соответствующий нервный механизм. Известно, что процессы, возникающие в так называемых ассоциативных зонах, воздействуют на функции воспринимающих систем. Экспериментальные данные говорят о том, что посредством этих путей осуществляется контроль над направленностью внимания со стороны подкорковых, а возможно, и корковых фильтров, или экранов, в пределах входных систем. Эти данпые получены в экспериментах с применением методики оценки скорости восстановления возбудимости во входных каналах (см. Pис. Участками, где, вероятнее всего, обнаруживается такое влияние, являются, конечно, те, где осуществляются тормозные взаимодействия нейронов, иначе говоря — «на входе». Эти взаимодействия образуют фильтры, через которые и на которые передаётся вводимая информация. Например, путём увеличения латерального торможения может быть усилен сенсорный контраст и замедлено восстановление системы (поскольку считается, что латеральное торможение реципрокно влияет на затухание; см. Pис. Замедление восстановления возбудимости в первичной зрительной системе действительно наблюдается при стимуляции нижневисочной коры (Spinelli and Pribram, 1966, Рис. — Поскольку нам нужен был более стабильный индикатор возбудимости, мы временно отказались от многократных предъявлений вспышки света. Мы полагали, что сетчатка является нестабильной зоной и что электрическая стимуляция более центрального участка зрительной системы посредством вживлённого электрода даст более надёжные результаты. Возможно, однако, "что за достижение стабильности нам пришлось бы заплатить деной утраты возможности оказывать влияние на возбудимость системы. И действительно, электрическая стимуляция нижневисочной коры не смогла оказать влияние на возбудимость, определяемую корковым электродом.
Эту дилемму разрешил наш сотрудник Л. Гербрандт посредством простого наблюдения. Он показал, что амплитуда ответов на электрическую стимуляцию зрительной системы зависит от уровня внимания обезьяны во время опыта. Когда обезьяну сажали в закрытый ящик, реакция была слабой. Когда же ящик открывали и обезьяна получала возможность оглядываться по сторонам, реакция усиливалась. Более того, раздражение нижне-височной коры могло превращать слабый ответ, полученный в закрытом ящике, в сильный, но оно не оказывало влияния на «ильный ответ. И наконец, используя величину зтого вызванного при помощи электрической стимуляции ответа в качестве контрольного, Гербрандт мог предсказать, повлияет ли раздражение лижневисочной коры в условиях закрытого ящика на функцию восстановления возбудимости зрительной системы или нет. В случаях, когда обезьяна была внимательна, ранее полученные нами.результаты не наблюдались. Когда же обезьяна начинала «скучать», клевать носом и так далее, влияние электрического раздражения на функцию восстановления было очевидным. В наших первоначальных экспериментах мы по заведённому порядку проделывали длительные ежедневные процедуры: парные вспышки, парные щелчки, комбинации щелчок — вспышка и вспышка — щелчок, вспышки по определённому шаблону и так далее, причём всё это предъявлялось в определённом порядке, изо дня в день, из недели в неделю. Не только обезьяны, но Эти эксперименты, вместе взятые, показывают, что результаты электрической стимуляции нижневисочной коры и результаты, получаемые, когда обезьяна «внимательна», сходны и что эти два процесса характеризуются высоким уровнем конвергенции на Таким образом, влияние височной доли на систему входа представляет собой частный случай и результат Фильтры признаков и механизмы опознанияМногочисленные исследования показали, что узнавание предполагает идентификацию признаков, различающих объекты. На этом основывается имитация этого процесса на ЭВМ при составлении программ. Открытие нейронов, чувствительных к определённым признакам, то есть способных избирательно реагировать на линии, наклонные под разным углом, движение, цвет и так далее, стало поэтому считаться решением проблемы узнавания. Не умаляя значения этого открытия, мы показали в предыдущих главах, что нельзя объяснить все проблемы восприятия наличием механизма детекции признаков. Позвольте мне развить эту точку зрения, высказав предположение, что проблема узнавания, зависящая от памяти, также останется неразрешённой, если мы будем основываться только на детекции признаков. Поэтому я разграничу детекцию признаков и их опознание. Детекторы признаков являются необходимым звеном нервного аппарата и не могут быть радикально модифицированы в результате накопления опыта, если они должны выполнять задачу выделения признаков. Поэтому детекторы признаков — это стабильные, «встроенные», врождённые элементы входных систем, которые проводят предварительную обработку сигналов, до того как они подвергнутся дальнейшей обработке. Напротив, механизм идентификации признаков, на котором основывается узнавание, должен по необходимости быть гибким и поддающимся модификациям в процессе приобретения опыта. Однако в силу непосредственности узнавания при предварительной обработке доходящей до мозга информации происходит одновременно как выделение признаков, так и процесс их опознания. Это участие двух разных процессов и привело к смешению этих двух механизмов Детекторы признаков воспринимаемых объектов играют довольно-таки ограниченную роль. Полагают, что они наряду с другими механизмами обеспечивают необходимый уровень, фон, на котором возникают другие, более подвижные конфигурации нервных событий. Они представляют собой постоянные элементы, «основу», на которой возникает узор микроструктурных изменений, чувствительных к влиянию приобретаемого опыта. В любой данный момент эта «основа» подвергает обработке воздействующие на неё нервные сигналы, то есть подготавливает их к переключению на последующие клеточные элементы. Процесс обработки не оказывает влияния на саму «основу», но добавляет к ней ещё один новый узор — ещё одну нить. Ряд таких механизмов, посредством которых опыт может оказывать влияние на нервную систему, был детально описан в главах II и XIV. Эти механизмы, Можно, пожалуй, считать, что нервный механизм, посредством которого осуществляется опознание, действует сходным образом. Обычно организованные элементы фильтра, колонки кортикальных клеток более или менее связаны между собой посредством элементов, чувствительных к направлению. Напомним, что в Известно, что макромолекулы типа липопротеинов и глюкоидов, образующие синаггхические и дендритные мембраны в микроструктурах мозга, подвержены таким изменениям. Всякий раз, когда нервный сигнал проходит через такую мембрану, эти изменения имеют тенденцию стабилизироваться, упорядочивая молекулярные структуры, пока длится сигнал
Такая система диполей, структур нейронных реестров имеег определённые преимущества, потому что она представляет собой, параллельно обрабатывающий механизм, связанный с содержанием. Элементы такой системы действуют одновременно, а непоследовательно; место хранения информации теряет своё значение. Предварительная обработка и модификация, основанные на опыте, происходят бок о бок и практически одновременно. Короче говоря, некоторые части механизма предварительной обработки изменяются в процессе этой обработки. Нервный механизм самоорганизуется. Подведём итог. В реальной жизни нелегко разграничить перцептивный процесс формирования Образа и познавательный процесс его опознания. Мы опознаем то, что ощущаем как значительное, почти в тот же момент, когда воспринимаем это посредством своих органов чувств. Но для целей научного анализа, особенно если нас интересует механизм, важно расчленить, как мы и сделали в этих главах, различные участвующие в этом процессы. Поскольку в итоге мы выясняем, что нервные механизмы этих процессов в действительности тесно взаимосвязаны, мы приходим к более полному пониманию того, каким образом возникает единство психологического процесса. Когда это единство ясно выражено, знак опознается или актуализируется, но эта актуализация (как и все действия) должна осуществляться последовательно. Любое мгновенное формирование знака поэтому не является полным — знак извлекает значение только из целостного Образа и только тогда может заменить его. Значимость
Нам осталось обсудить последний вопрос: как некоторые аспекты Образов обретают смысл, становятся значимыми? В экспериментах по выработке дифференцировок у обезьян какое-либо изображение получает смысл, становится знаком в результате поведения самой обезьяны. При выборе используется нервный механизм, необходимый для совершения действия. Тот факт, что проводящие пути от нижневисочной области коры, которая оказывает влияние на зрительное внимание, проходят через двигательные отделы мозга, обеспечивает структурную базу для взаимодеиствия двигательных и сенсорных процессов и для влияния Образов результата на Образы восприятия. Поэтому значение данных сигналов создаётся посредством действия. Именно, этот активный процесс образования знака порождает значение: он является результатом перцептивного научения посредством подкрепления. Оказывается, что значимость сигналов зависит от механизма, который привлекает к ним внимание, от подкреплений и альтернатив (см. Pис. Работа этого механизма весьма напоминает укорочение фокусного расстояния транслокаторного фотообъектива — при этом в фокусированное поле зрения попадает большая площадь. Были получены прямые доказательства наличия такого механизма у человека. Измерения рецептивных полей отдельных элементов в зрительной коре человека показывают, что константность восприятия величины достигается здесь при помощи процесса, поддерживающего постоянный угол относительно точки фиксации взора в геометрическом центре рецептивного поля (Marg and Adams, 1970). Так, когда я хочу рассмотреть большую часть какой-либо сцены, я укорачиваю фокусное расстояние. Парадоксально, что, когда затем меня начинает интересовать тот или иной аспект этой сцены, соответствующие признаки воспринимаемого выделяются и становятся свободными от влияния побочных деталей Когда же, напротив, я намереваюсь уточнить Это сужение фокуса в зависимости от контекста представляет собой другой аспект проблемы значимости. В РезюмеСвоими достижениями человек в большой степени обязан своим уникальным коммуникационным способностям. Эти способности развиваются, когда в восприятие и переживание включаются моторные механизмы мозга. Возникающие в результате этого кодирующие процессы создают знаки и символы. Знаки создаются и опознаются тогда, когда моторные механизмы действуют на структуру нейрональных соединений, структуру, образуемую поступающей информацией, а символы — когда моторные механизмы воздействуют на сенсорные процессы в центральной части ствола мозга (Различение символов и знаков, предлагаемое автором, нуждается, безусловно, в дальнейшем уточнении. — | |||||||||||||||||||||||||||||
Оглавление | |||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| |||||||||||||||||||||||||||||