«На более высокой стадии понимания поведения организма «исторические» переменные заменяются «причинными». Если мы сможем наблюдать текущее состояние организма… и, если мы сможем непосредственно создавать или изменять это состояние, то мы сможем использовать зги знания для контроля над поведением». Б. Ф. Скиккер. «Вместо постоянного повторения того, что с помощью подкрепления можно достигнуть контроля над поведением, я хотел бы подчеркнуть, что подкрепление может привести к… (достижению результата, основанному на) компетентности организма. И Дж. Миллер. Единство всех «последовательных реакций является краеугольным камнем формирования индивидуума, сохранение целостности которого и составляет особую задачу нервной системы» (Шеррингтон, 1969. —
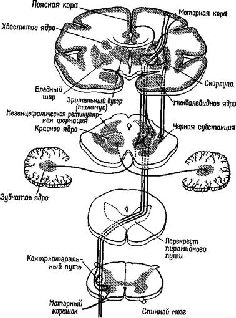
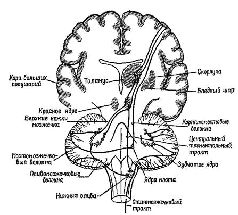
В главе III, IV, VII и VIII обсуждалось формирование программ, состояний в мозгу как функции рецепторов и системы, перерабатывающей поступающую информацию. Мы видели, что формирование подобных программ зависит от образования некой конфигурации из ряда однородных элементов, различные наборы которых размещаются в ней в том порядке, в котором они появляются в этой конфигурации, причём каждый из элементов конфигурации в отдельности представляет всю структуру в целом. В качестве примера служил «разговор» с компьютером, обладающим двоичным контролем, то есть таким компьютером, переключатели которого могут находиться только в двух позициях. Такой компьютер с трудом различает последовательности типа ААВВВА и АВВААВ, но кодирование этих последовательностей в виде рядов восьмеричной или десятичной системы делает общение с данным компьютером возможным. В равной степени важно — с точки зрения проблем, поставленных в этой главе, — что декодирование восьмеричной (или десятичной) системы в двоичную должно осуществляться прежде, чем оператор будет работать с компьютером. Операция декодирования противоположна той, которая требовалась при кодировании; теперь набор различных элементов должен быть превращён в более элементарную структуру, состоящую из однородных элементов. Это и есть проблема последовательной организации поведения, иначе говоря, — проблема единства действия во времени. Модификация структуры должна осуществляться как по пространственным, так и по временным характеристикам, что и создаёт соответствующий код. Подобно тому как симфония композитора должна одновременно декодироваться в движения смычка и его остановки, определённая конфигурация в мозгу должна быть превращена в организованные по пространственному и временному принципам структуры нервных импульсов, которые регулируют степень сомкнутости голосовых связок, чтобы изменять прохождение через них воздуха. Даже при простом сгибании передней лапы собаки набор сигналов должен генерироваться таким образом, чтобы во время реципрокно связанных сокращений и расслаблений сгибателей и разгибателей все остальные части тела оставались неподвижными. Рассматривая с этой точки зрения организацию поведения, его последовательный характер следует понимать не как соединение движений в цепи, а как их дифференциацию, процесс декодирования ранее сформированной пространственной конфигурации. Таким образом, во многих отношениях проблема поведения является обратной стороной проблемы Образа. Образ — это отражение внутри организма его окружения, а поведенческий акт — это отражение в окружающей среде того, что имеется внутри организма. Поведенческие акты — это достижение результатов; так, например, построение гнёзда или написание книги совершается во внешнем мире, как оно было запланировано организмом, то есть соответствует его намерению или установке. То, как организуется поведенческий акт, и составляет предмет этого раздела нашей книги. Вернёмся ещё раз к модели продуцирования слов, о которой мы говорили в начале этой книги. Предложение «Я вас люблю», произнесённое машиной, есть некий акт. Правда, здесь смоделирован только периферический голосовой акт, иными словами, электронная машина заменила собой уникальный голосовой аппарат человека. У человека настройка голосового инструмента должна осуществляться с помощью мозга. Тем не менее этот инструмент действует таким образом, что генерация даже такой сложной активности, как произнесение предложения, может осуществляться лишь посредством модификации двух мозговых процессов. Каким же образом тогда осуществляются эти модификации? Я считаю, что организация действия есть в значительной мере управление рецепторными механизмами, содержащимися в сокращающейся ткани (в мышечных веретенах) или в сухожилиях, которые прикрепляют мышцы к костям и сочленениям. Эти рецепторы реагируют не только на сокращения мышц независимо от того, вызываются ли они внешними силами или нервными импульсами, возникающими в мозгу (через систему эфферентных волокон большого диаметра, известных под названием альфа-системы), но также и на возбуждения, приходящие к ним непосредственно через гамма-эфферентную систему волокон. Следовательно, непосредственная организация движения прямо связана с процессами, в которые вовлечены рецепторы, афферентные пути, идущие от этих рецепторов к спинному мозгу, и эффекторные пути, идущие от спинного мозга к сокращающимся мышцам и рецепторам (см. Pис. Всякий раз, когда этот процесс завершается стабильным ответом на стимуляцию, мы отождествляем его с рефлексом. В этой главе я впервые остановлюсь на некоторых интересных проблемах, касающихся осуществления рефлекса, который возникает благодаря наличию центрального контроля над рецепторами. Затем я перейду к детальному анализу мозговых механизмов управления функциями мышечных рецепторов. Сервоконтроль мышечных сокращенийВ связи с существованием непосредственного контроля со стороны центральной нервной системы над рецепторными процессами посредством гамма-системы афферентных волокон, нейронную организацию рефлекса нельзя больше рассматривать как дугу или простую последовательность «стимул — реакция». Существует множество данных, свидетельствующих о том, что и сами рецепторы контролируются центральной нервной системой — особенно те из них, которые воспринимают мышечные сокращения. Этот центральный контролирующий механизм действует как система обратных связей и процессов предвосхищения, что, по существу, и определяет корригирующие возможности сервомеханизма. Получено так много доказательств справедливости этого положения, что Р. Гранит, один из первых исследователей как моторных, так и сенсорных механизмов, вынужден был недавно заявить следующее: «… Что касается мотонейронов, то основные проблемы связаны не с рефлексами, хотя рефлексы всегда были и всё ещё остаются полезным средством анализа, необходимым в данной области знания. Основные проблемы касаются настройки или установки различных механизмов, посредством которых управляются мотонейроны — рефлекторно или Относительно сервоконтроля движений многое стало известно после эксперимента, проведённого в духе нейропсихологических исследований и давшего парадоксальные результаты.
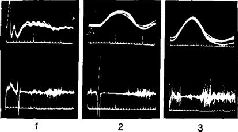
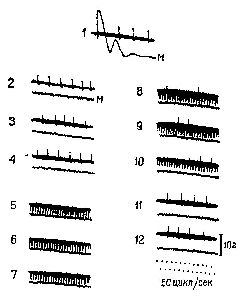
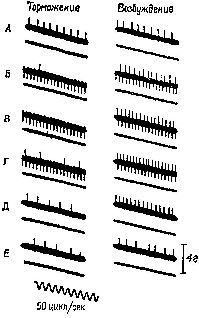
«Если мышца длительное время находится в состоянии сокращения под влиянием разрядов моторных импульсов, идущих от спинного мозга, удар тока, нанесённый на моторный нерв, приведёт к возникновению судорожных всплесков на тензозаписи. Во время судорожного сокращения наблюдается прекращение разрядов моторных импульсов, идущих к мышце (см. Pис. ХII–3). Это — период молчания. Это — рефлекторный акт (обсуждение этого вопроса см. Merton, 1951), который возникает, Надо отметить, что отрицательная обратная связь не только нейтрализует эффекты изменений нагрузки или интенсивности добавочного потока импульсов, но делает работу мышц независимой от утомления или изменения синаптической возбудимости, точно так же, как усилитель, сконструированный по принципу обратной связи, не чувствителен к изменениям интенсивности поступающего тока или к напряжению электронных ламп. Стабильность — вот единственное требование, предъявляемое к системе обратных связей: в усилителе это сеть обратных связей со стабильным сопротивлением, в мышцах же это неутомляемые сенсорные органы. Как известно из работ Мэтгьюза, мышечные веретена как раз и обладают этими свойствами… Возвращаясь к проприоцептивному механизму периода молчания, следует сказать, что мышечные, веретена прекращают свои разряды во время судороги потому, что они соединены параллельно с главными мышечными волокнами. При сокращении мышцы натяжение веретён уменьшается и они не посылают больше разрядов. Это «параллельное» соединение веретён и является ключом ко всей теории. Важная роль этого механизма состоит в том, что он даёт возможность веретенам посылать сигнальную информацию о длине мышц, тогда как если бы веретена были соединены последовательно, они могли бы отвечать только на их напряжение. Таким механизмом, как серворефлекс растяжения, который столь очевидно обеспечивает поддержание определённой длины мышц, а не их напряжения, должны обязательно обладать рецепторы, сигнализирующие о длине мышц» (Merton, 1955. — Существуют, конечно, и другие рецепторно-эффекторные связи, контролирующие сокращения мышц. Есть рецепторы в сухожилиях и суставах — нельзя не учитывать также и растяжение кожи вокруг мышцы и суставов как источник информации для контроля над движениями. Хотя о, рамма-системе известно больше, чем о других механизмах контроля движений, иннервация, поступающая от мышечных веретён, является, по всей вероятности, основным механизмом, на котором надстраиваются другие контролирующие механизмы (см. Pис. ХII–4). Как было установлено, гамма-волокна бывают двух типов. Один тип реагирующих волокон обнаруживает постоянную частоту разрядов, если мышца сохраняет постоянную длину. Другой тип, напротив, обнаруживает уменьшение частоты разрядов во время завершения динамической фазы растяжения. Мэттьюз (1964) в обширном обзоре, посвящённом этой теме, приходит к заключению, что такая двойная гамма-иннервация каждого мышечного веретена «обеспечивает относительно независимый контроль над общим режимом работы и отключением сервосистемы», поскольку каждый тип контроля раздельно управляется с помощью центральной стимуляции (см. Pис. ХII–5).
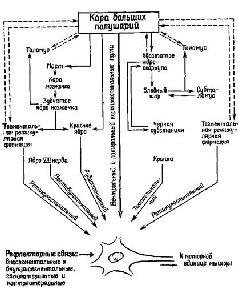
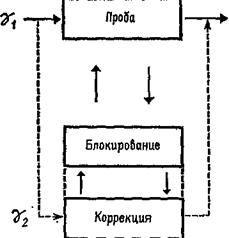
Этот анализ важен с двух точек зрения. Во-вторых, управление рефлекторным сервомеханизмом легко представить себе, поскольку этот процесс разыгрывается на уровне рецепторов. Следовательно, нет необходимости в специальном устройстве типа клавиатуры фортепиано, с помощью которого импульсы, идущие от мозга, передавали бы мышцам сообщения; дело вовсе не в том, чтобы осуществить сначала одно, а потом другое сокращение, изменить длину того или иного из мышечных волокон. Существующее состояние сокращения мышечного волокна предопределяет вариативное состояние его рецепторов. Набор нервных импульсов взаимодействует с этим фоновым состоянием, изменяя режим работы мышечной сервосистемы и таким образом регулируя мышечное сокращение. Мозг всё время должен посылать сигналы, организованные по пространственному и временному принципам, однако сообщения, передаваемые этими сигналами, не должны быть закодированы таким образом, чтобы прямо соответствовать сокращению или расслаблению мышцы, которое эти сообщения вызывают, поскольку эта информация уже содержится в вариативном состоянии рецептора. В следующей главе мы детально проанализируем эти сообщения, которые должны содержать подобный код. Подведём итоги. Нейрофизиологические данные свидетельствуют о существовании прямых анатомических афферентных путей, идущих от спинного мозга к мышечным рецепторам. Наиболее фундаментальным фактом является то, что основным принципом организации даже самого простого рефлекса является не дуга: стимул — центральная нервная система — реакция, а замкнутая сервосистема в виде последовательности: проба (test) — операция (operate) — проба (test) — результат (exit) (Т—О—Т—Е). Когда с помощью центральной нервной системы рефлексы объединяются в Сложные движения, эта интеграция не может быть эффективной при посылке серий сигналов непосредственно или исключительно сокращающимся мышцам, вызывая в них мелодии, как на клавиатуре фортепиано. Подобные сигналы только бы нарушали сервопроцессы. Чтобы предотвратить эти нарушения, наборы сигналов должны передаваться либо только мышечным рецепторам, либо одновременно с другими сигналами адресоваться непосредственно к мышечным волокнам. Таким образом, интегрированное движение в основном зависит от режима работы, настройки мышечных рецепторов. Подготовка к ответуТеория, излагаемая в этой книге, вполне согласуется с взглядами Ф. Меттлера (1967). Последний считает, Далее, Меттлер признает существование «ассоциированных массивных движений (предмет данного раздела), на фоне которых вырисовываются отдельные детали…» Массивные процессы регуляции мышечного тонуса осуществляются базальными ганглиями, самыми передними структурами ствола мозга. Наконец, «на этом фоне статических и глобально действующих сил кора мозга ответственна за точность и интенсивность движений, з также за изменение скорости уже начавшегося движения и той скорости, с которой это движение усиливается и прекращается». Статическое состояние функций мышцы называется её тонусом. Однако тонус — это не величина спастичности или вялости сократительной ткани. Это, скорее, состояние готовности всего нейромышечного аппарата к действию, предварительное условие всякого действия.
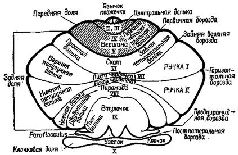
Центральными структурами с замкнутыми системами связей, участвующими в регуляции тонической мышечной активности, являются базальные ганглии (см. Pис. Регуляция мышечной активности с помощью замкнутых кольцевых связей базальных ганглиев и передних отделов мозжечка была предметом большого числа исследований, однако в этом вопросе до сих пор нет полной ясности.
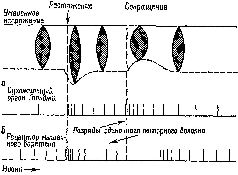
Факт произвольного движенияСервоконтроль имеет и другой аспект. Иногда сервомеханизм не только регулирует движение «на выходе», но и создаёт предварительную установку к следующему этапу, то есть обеспечивает то предшествующее двигательной активности состояние, которое нуждается в стабилизирующих влияниях сервомеханизма. Эти процессы предвосхищения (см. Pис. «Что же происходит во время произвольного сокращения? Легко осуществить произвольное сокращение мышц при устойчивом уровне двигательной активности; эти движения совершаются даже быстрее, чем самые быстрые судорожные сокращения. Скорость возникновения небольшого судорожного сокращения мышц лишь в малой степени соответствует скорости произвольного усиления. Здесь возникает очень интересная проблема: если во время такого усиленного сокращения мышц сервомеханизм будет продолжать действовать так же, как прежде, то он явно будет способствовать противоположному эффекту. Сразу же после начала произвольного сокращения мышечное веретено будет укорачиваться и возникающее в результате этого прекращение возбуждающих афферентных импульсов будет интенсивно подавлять мотонейроны… как если бы сокращение мышцы вызывалось небольшой стимуляцией двигательного нерва. Это значит, что дополнительные моторные разряды, возникающие при возбуждении мотонейронов спинного мозга, должны вызывать гот же самый эффект в замкнутой сервосистеме, что и импульсы, возникающие вокруг этой системы в их аксонах под влиянием электрического тока. В любом случае сервомеханизм должен энергично действовать, чтобы прекратить эти механические эффекты. Если эти выводы из экспериментов верны, то это означает, что во время произвольного сокращения следует преодолеть или устранить сопротивление, оказываемое сервосистемой… необходимость возбуждения основных мотонейронов, связанных с интрафузальными мышечными веретенами посредством нисходчщих импульсов, идущих из спинного мозга, исчезает. Возбуждение целиком поступает к небольшим мотонейронам (рецепторам мышечных веретён), н сервомеханизм запускает затем в действие основные мышцы. Таким образом, в произвольном сокращении мышцы участвует тот самый механизм, который обеспечивает постоянное фоновое сокращение, а именно: сервомеханизм, поддерживающий в мышце такой же ритм, как Эта гипотеза в известной мере подтверждается экспериментами, недавно проведёнными на нервно-мышечных приборах, которыми снабжена грудпая мышца. Хант (1951), в частности, наблюдал во время рефлекторного сокращения этой мышцы разряды моторных импульсов, посылаемых к интрафузальным мышцам (рецепторам веретён), предвосхищающие появление активности в основных мотонейронах» (Merton, 1953. — Таким образом, произвольные движения, как правило, начинаются с активации интрафу зальных мышечных веретён. Исключением из этого правила является тот случай, когда движения возникают неожиданно. Вследствие сравнительно малого диаметра гамма-волокон и большой длины всего пути скорость проведения слишком мала, чтобы объяснить внезапные мышечные сокращения. Однако подобные движения относительно просты и гамма-система быстро включается в контроль над этими движениями. Следовательно, нельзя полностью игнорировать прямой путь к основным мотонейронам. Ограниченный объём предварительной информации, которая программирует процесс «сокращения — расслабления», может быть передан по этим путям, если интервал между началом и окончанием процесса будет достаточен для того, чтобы могла включиться в действие гамма-сервосистема. Работы Дж. Бермана и его сотрудников (Taub, Bacon and Berman, 1965) установили как верхнюю, так и нижнюю границы такого рода предпрограммирования движений. Берман использовал в эксперименте обезьяну, у которой спинной мозг ла всем его протяжении был полностью деаффектирован посредством удаления дорсальных корешков. Эта операция вызывает нарушение замкнутого сервомеханизма, обслуживающего движения. Однако после деафферентации у этих обезьян всё же сохранялись условные реакции (сгибания конечностей). У человека же подобная деафферентация, широко практикуемая в настоящее время, чтобы снять боли и другие сенсорные нарушения, приводит к серьёзным расстройствам автоматизированных движений, выполняемых соответствующей конечностью. Итак, управление рефлекторными и интегрированными произвольными движениями независимо от того, вызываются ли они внешними или внутренними факторами, осуществляется в основном посредством настройки рецепторов мышечных веретён, включённых в гамма-сервосистему. Центральный контроль над движениями сводится, следовательно, к центральному контролю над рецепторными процессами. Эту проблему мы уже обсуждали во второй части, более подробно рассмотрим её в четвёртой части книги. Теперь мы сформулируем парадокс, который не укладывается в рамки узкого бихевиоризма: даже относительно простые, доступные для наблюдения двигательные механизмы связаны с рецепторными процессами, регулируемыми центральной нервной системой. Быстродействующие механизмы предвосхищенияПризнав факт существования произвольных движений, мы должны ответить на вопрос: как же они осуществляются? Ответ на этот вопрос можно дать на основе анализа процессов предвосхищения, с одной стороны, и раскрытия сущности предпрограммирующих процессов, подготавливающих результаты ряда движений прежде, чем они будут реализованы, — с другой. Вспомним, что в
Как уже отмечалось выше, передняя часть мозжечка развилась при переходе морских позвоночных к наземному образу жизни. До этого момента мозжечок функционировал главным образом как дополнительный вестибулярный контролирующий механизм. Рыбы поддерживают ориентацию в соответствии с осью своего тела; их движения начинаются от этой основной линии. Амфибия и другие земноводные имеют дополнительную основную линию, это земля под ними, «низ», определяемый посредством гравитационных сил. Поэтому для организма становятся полезными новые вертикальные ориентиры позы и вследствие этого развивается передняя часть мозжечка. Наконец, приматы в известной мере освобождаются от гравитационной основной линии благодаря развитию двигательных навыков и приспособляются к древесному образу жизни и прямой позе. Теперь полушария мозжечка становятся доминирующими. У человека повреждение этих отделов мозга ведёт к нарушению координации движений, к грубым промахам при слежении за целью
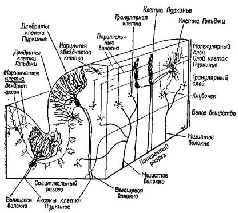
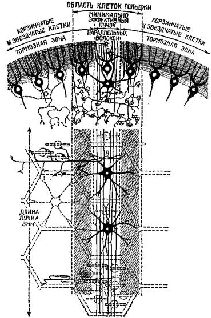
К счастью для нейрофизиологии, полушария мозжечка имеют удивительно гомогенное строение (см. Pис. Сущность этих процессов сравнения становится особенно интересной в свете последних детальных анатомических и физиологических данных (см. Pис.
Быстрое стирание содержания записи предполагает, что последовательность операций сличения выполняется тоже довольно быстро. Эта особенность системы выходит за рамки обычного сервомеханизма обратных связей, в котором для длительного поддержания устойчивого состояния требуется постоянная информация о результатах движения. Т. Рач в своей работе, опубликованной в «Экспериментальной психологии» (1951), правильно сформулировал эту проблему, причём значительно раньше, чем были собраны соответствующие факты.
Основная функция мозжечка, связанная с объёмом регистрации, заключается в том, чтобы составить схему движения без детальной его разработки. В действительности такое кратковременное состояние, которое аналогично тому, что создаётся порой больших полушарий, формируется на фоне спонтанной активности.
Подробнее я остановлюсь на этом приборе и на его аналоге — в коре больших полушарий в РезюмеПроблема мозговой организации поведения во многих отношениях является обратной стороной формирования Образа (восприятий и чувств). Вследствие повсеместного существования в нервной системе механизма обратной связи (и механизма предвосхищения) контроль над движениями достигается не непосредственно с помощью сигналов, адресующихся к сокращающимся мышечным волокнам, а посредством сигналов, адресующихся к рецепторам, которые регулируют мышечное сокращение. Таким образом, мышечные рецепторы становятся частью гармоничной системы (сервомеханизма Т—О—Т—Е), назначением которой является регуляция мышечных реакций в ответ яа изменения внешней среды. Короче говоря, нервная регуляция поведения осуществляется посредством воздействия на рещепторные процессы, а не через непосредственный контроль мышечных сокращений. | |||||||||||||||||||||||||
Оглавление | |||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| |||||||||||||||||||||||||