Бихевиоризм: стимулы, ответы и подкреплениеВ предыдущих главах мы проводили различие между движением и действием. Движения, то есть организованные мышечные сокращения, создают набор поз, двигательный контекст или матрицы, на основе которых формируются акты поведения. В нервный механизм, контролирующий действие, включена информация, поступающая из внешней среды, которая создаёт нечто вроде силовых полей, но в ней не содержатся сведения о длине той или другой мышцы Неожиданно оказалось, что вопрос, поставленный нейрофизиологом, входит в сферу интересов психолога. Специфика этих внешних условий, определяющих действие, и есть то, что всегда изучает бихевиорист, пытающийся построить свою концепцию на анализе организации внешней среды, значимые связи которой получают объяснение через поведение организма. Бихевиористы правильно считают, что стимулы, ответы и подкрепление получают своё значение на основании внешних, а не внутренних условий существования организма, хотя необходимо учитывать и поведение организма, без которого картина останется неполной. Почему тогда психологам не ограничиться только «изучением доведения»? Отношения между стимулом, ответом и подкреплением сложнее, чем это представляется бихевиористу. Хотя центр интересов бихевиориста меняется, все бихевиористы — за небольшим исключением — представляют себе эту связь как прямую последовательность. Стимул -› ответ - подкрепление является той самой последовательностью, которая видна наблюдателю. Однако то, что эта очевидная последовательность есть результат более сложных взаимоотношений, стало ясно только тогда, когда обнаружилось, что мозг обеспечивает этот порядок.
Возьмём, например, тот факт, что рефлекс организуется не па схеме дуги S — R, a с помощью сервосистемы. Этот факт оказал важное влияние на понимание сущности стимула и ответа. Существование механизмов обратных и предваряющих связей усложняет всем известные представления Ньютона и Шеррингтона о линейной связи стимула и реакции. В результате перед бихевиористами возникли две возможности: либо игнорировать внутреннюю сложность этой системы, либо пытаться объяснить её и дать трактовку тем изменениям, которые претерпевает эта система. В большинстве случаев бихевиористы игнорировали сложность системы. Однако они не могут уйти от того, что стимулы сами находятся под воздействием возникающих ответов Согласно такому логически и неврологически обоснованному пониманию взаимоотношений между
Возможно, наиболее интересным для анализа этой проблемы «позиций бихевиоризма является объективность, достигнутая в определении подкрепления, когда схема S-› R, или концепция флекторной дуги, заменяется теорией сервомеханизмов. Агентом, запускающим сервомеханизм, является проба на соответствие или несоответствие между внешними образами, текущим «состоянием» и характером энергии, воздействующей на это состояние. Тйким образом, то, что психологи называют ближайшим («проксимальным») стимулом, зависит от состояния системы, открытой для ввода информации, то есть от «установок» и «ожиданий» — по терминологии психологии познавательных процессов. Другие рассуждения ставят подкрепляющие свойства среды в зависимость от совпадения условий, которые вызывают данное поведение, с теми условиями, которые оно само вызывает. В этой главе, а также в Совпадение и контекстПомимо очень большого вклада в технику эксперимента, бихевиористский подход к изучению поведения значительно обогатил науку своим анализом модификации поведения. Однако объект исследования бихевиоризма ограничен. Бихевиоризм изучал только те изменения поведения, которые непосредственно вызывались стимулами, исходящими от экспериментатора и побуждающими животное к действию. Бихевиористы-экспериментаторы чаще всего не принимали во внимание изменений в поведении, возникающих в результате участия эмоциональных процессов в механизме самоконтроля. Они не придавали должного значения и структурным законам, лежащим в основе изменчивости поведения (Pribram, 1970). Результаты их экспериментов, проведённых главным образом на животных, обедняли теорию бихевиоризма, которая в настоящее время нуждается в уточнении за счёт фактов, связанных с деятельностью мозга. Это и есть тот узкий круг вопросов, которые мы будем обсуждать. С позиций бихевиоризма, изменения поведения происходят благодаря подкреплению. А что же такое подкрепление? Перед последовательными сторонниками бихевиоризма подобный вопрос не возникает: их интересует только описание тех явлений окружающей среды, которые вызывают эффект подкрепления. (Skinner, 1969). Однако следует отметить, что, даже согласно такому узкому определению, подкрепление должно быть Согласно другой позиции, которой придерживаются сторонники теории влечений и установок, для достижения определённых результатов в научении необходимо изменение внутреннего состояния организма. Как мы увидим дальше, «одна лишь ассоциативная» концепция оказывается несостоятельной для анализа того, что же происходит в мозгу при научении. Сторонники теории ассоциаций подразделяются на две категории: одни из них предпочитают язык ответов, другие — язык стимулов. И те и другие полагают, что поведение определяется только набором стимулов, организованных по принципу вероятности, причём значение этих стимулов определяется их вероятностью (Estes, 1959). Сторонники теории стимулов полагают, что значение стимулов определяется условиями эксперимента, то есть главным образом вычисленными отношениями между стимулами. Сторонники теории ответов считают, что стимулы приобретают значение благодаря многократно осуществляющимся актам поведения, вследствие чего само поведение становится «саморегулируемым» (Guthrie, 1942).
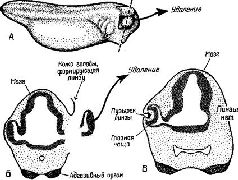
Подход к проблеме регуляции поведения с позиций теории простого совпадения стимулов и подкрепления сталкивается с рядом фундаментальных вопросов: 1) может ли происходить научение при отсутствии действия (то есть подкреплённого поведения) и, если это возможно; 2) что способствует научению, когда животное выполняет действие? Мы уже знаем, что научение может осуществляться и без действия. Когда в относительно постоянной ситуации организму многократно предъявляются экспериментально контролируемые сигналы, происходит привыкание организма. В 1960-х годах нейрофизиологи много занимались проблемой привыкания. В результате их исследований наше понимание этого процесса радикально изменилось. Рассмотрим кратко очень важный эксперимент, выполненный Таким образом, рассуждал Соколов, привыкание не может быть просто Как мы уже знаем, множество фактов подтверждает существование самоадаптирующегося механизма, своего рода фильтра, через который проходят поступающие сигналы Биологическая концепция, согласно которой ткань должна быть «компетентной», для того чтобы дифференцироваться, является результатом ряда работ, авторы которых пытались наглядно показать, как возникает и контролируется процесс дифференциации ткани, то есть как начинается её развитие. Толчком к развитию служит, в сущности, химическое взаимодействие между «компетентной» тканью и организующими воздействиями среды, которые направляют развитие этой ткани. Классическим примером является один из ранних экспериментов, в котором развитие хрусталика определялось глазным пузырьком. Контакт между этим пузырьком и внешним слоем эпидермы стимулирует пузырёк к образованию хрусталика на месте контакта Если глазной пузырёк удалён, эпидерма не способна превратиться в хрусталик (см. Pис. Этот эксперимент поставил целый ряд проблем, из которых родилось новое направление исследований в экспериментальной эмбриологии, имевшее поразительное сходство с направлением исследований в экспериментальной психологии и этологии. Сначала предполагалось, что индуктор действует только как пусковой механизм (триггер); так, в классическом примере считалось, что эпителий головного конца уже «предрасположен» к образованию хрусталика и требуется только сигнал к старту Эта концепция индуктора была опровергнута двумя видами данных. Кроме этих фактов, существуют другие, которые показывают, что реагирующая ткань должна быть «готова» или «компетентна», то есть она должна находиться в соответствующем состоянии реактивности, чтобы индукция стала эффективной. Например, ткань, процесс превращения которой в различные специализированные структуры уже «запущен», совершенно не отвечает на индукцию. Далее, было обнаружено, что индукторы действуют независимо от принадлежности животного к тому или другому виду Один и тот же индуктор может быть эффективным по отношению к ткани, принадлежащей животным различных видов, родов и даже отрядов Следовательно, можно предположить, что в основе действия индукторов лежат химические свойства, общие для многих организмов (мы будем подробнее говорить об этом дальше). В результате большого числа химических экспериментов постепенно утвердилось мнение, что рибонуклеиновая кислота (РНК) является наиболее вероятным, а возможно, и единственным веществом, которое ответственно за эффект индукции (см. Niu, 1959), хотя и рибонуклеопротеины, и стероиды не могут быть полностью исключены из этого процесса. В большинстве случаев рибонуклеаза разрушает эффект индукции, хотя остаётся нерешённым вопрос, оказывает ли рибонуклеаза какие-либо иные воздействия на индуцируемую ткань, в которой нарушается процесс дифференциации. Однако более прямые доказательства роли РНК в процессе индукции были получены в опытах с демонстрацией эффекта индукции РНК, выделенной из различных органов. Было показано и то, что выделенная из различных органов РНК способна стимулировать реципиентную ткань к тому, чтобы она превращалась в различные специфические структуры. Эти эксперименты свидетельствуют о том, что в организме существует много видов РНК, каждый из которых имеет свои специфические функции. Итак, сторонники теории ассоциаций считают, что изменения поведения возникают всякий раз, когда события (стимулы) происходят одновременно или если такие одновременно происходящие стимулы (события) возникают благодаря ответам организма. Факты привыкания показывают, что «стимулы» с позиций теории ассоциаций должны рассматриваться как «проксимальные»; воздействие становится стимулом вследствие частичного совпадения «входной» информации с состоянием центров, которое в свою очередь формируется благодаря предшествующим частичным совпадениям центральных состояний и информации, поступающей от внешних воздействий. Таким образом, в любой отрезок времени центральные механизмы должны быть «компетентны», готовы к тому, чтобы обеспечить фон, на котором возникают стимулы. Следовательно, связь стимулов нужно рассматривать не как некую неопределённую, случайную, вероятную «ассоциацию» а как биологически детерминированный процесс, Который организует взаимоотношения контекста (фона) и воздействия стимула. Если это так, то теория совпадения стимулов и теория ожидания сближаются с точки зрения процессов, происходящих в центральной нервной системе. Таким образом, простого случайного сочетания, как такового, как фактора научения, — не существует, и эта концепция должна быть заменена биологической концепцией, основанной на принципе «компетентности»; наборы воздействий-стимулов (по существу, врождённых, хотя они и модифицируются в результате опыта) кодируются в центрах, которые приобретают «компетентность» по отношению к последующим наборам воздействий, создаваемых стимулами. Этот процесс модификации поведения напоминает процесс эмбриогенеза, последний определяется унаследованными, врождёнными свойствами, пробуждаемыми к жизни стимулирующей функцией среды, в которой растут клетки, Стимулирующая функция сама по себе специфична, но эта специфичность несколько иная, чем генетический потенциал. Генетическая «компетентность» индивидуально специфична (она специфична для каждого вида, рода, отряда). Наследственные факторы предполагают общность прошлого с будущим, допуская определённые вариации внутри каждого отдельного поколения. Индукторы, напротив, неспецифичны по отношению к индивидам, видам и так далее, поскольку существуют относительно простые химические агенты — РНК, общие для всех живых организмов. Таким образом, индукторы обеспечивают ту присущую всему живому общность, которая открывает возможность модификаций механизмов «компетентности» в отдельных поколениях в соответствии с требованиями момента. Смежность и последовательностьВторой вопрос, который возникал при бихевиористском подходе к проблеме, был следующим: что способствует научению, когда совершается действие? Можно сформулировать теперь этот вопрос иначе и спросить, происходит ли привыкание только при повторении стимулов или этот принцип применим к более широким проявлениям поведения. Хотя мы ещё не можем дать точный ответ на этот вопрос — для этого необходимы новые исследования, — было высказано предположение, что процесс привыкания и угашение поведения имеют общие факторы. Например, Д. Примак (Premack and Collier, 1962), анализируя случаи ненодкрепления, отражающиеся на вероятности ответов, счёл нужным заявить следующее: «Существует по крайней мере ряд данных о безусловных ответах, которые не сохраняются полностью при их повторном вызывании. Хотя эта проблема мало изучена, в одном из немногих имеющих отношение к данному вопросу исследований Доджа (1927) сообщается о небольшом, но явно необратимом уменьшении величины и увеличении латентных периодов даже такого безусловного рефлекса, как коленный. Далее, в биологической литературе о привыкании содержатся описания нескольких случаев, когда безусловные ответы уменьшались по мере повторения, а их латентные периоды увеличивались… Возникает вопрос… не имеет ли этот факт некоторого несомненного уменьшения безусловных рефлексов более широкое значение для понимания поведения, чем это принято считать» ( Только признав, что поведение регулируется механизмом, сходным с тем, который контролирует сенсорные процессы, можно, Подводя итоги проблемам, поставленным сторонниками теории ассоциаций или совпадения стимулов с подкреплением (ассоциаций по смежности), следует сказать, что ни теория «отбора стимулов», ни теория «саморегуляции поведения» не могут дать адекватного ответа на вопрос о модификации поведения, вероятно, потому, что эти теории не принимают во внимание важность организации процесса научения во времени. Организмы реагируют не на любые, одновременно происходящие смежные явления. Их поведение направляется предварительно сформированной «компетентностью» мозга, организующей стимулы и включающей в действие лишь те, которые важны для поведения. Таким образом, стимулы представляют собой воздействия, детерминированные нервной системой и «отобранные» на основании центральной «компетентности» организма (или его нервной «установки»), которая в свою очередь детерминирована предшествующим опытом и другими центральными процессами. Поведение организма направляется не только стимулами, но и посредством самоадаптирующегося фильтрующего механизма, который в определённых случаях сам индуцирует стимулы (то есть вызывает ориентировочную реакцию). Это происходит в тех случаях, когда результат поведения лишь частично совпадает с центральной «компетентностью», которая и программирует поведение. В этих условиях как раз и имеет место подкрепление и поведение становится саморегулируемым. Интересный эксперимент, проведённый Э. Джоном (John and Morgades, 1969), показал, что организм отвечает должным образом не только в силу того, что воздействия происходят одновременно. Три кошки были обучены нажимать на один рычаг при вспышках света частотой 2 кол/сек, и на другой, когда частота вспышек была равна 8 кол/сек. Регистрировалась электрическая активность в латеральном коленчатом теле — эвене зрительной системы. Было показано, что, когда животное правильно выполняло задание на различение вспышек, возникающие электрические ответы чётко различались по форме волн. У четвёртого необученного (контрольного) животного подобных различий в форме вызванных ответов не наблюдалось. Когда кошка ошибалась, то есть «огда давался неверный двигательный ответ при каком-либо сигнале, исчезало также и различие в форме электрических колебаний на этот сигнал. Джон пришёл к выводу, что за наблюдаемые различия, коррелирующие с правильными ответами, должны быть ответственны определённого рода динамические процессы (с точки зрения концепции, развиваемой в этой книге, «У тренированных в разной степени животных форма вызванных потенциалов, возникающих под каждым электродом в ответ на два различных стимула, отчётливо различается. Эти различия в вызванных ответах наблюдались при регистрации реагирующих ансамблей клеток в пределах ограниченных областей мозга. Таким образом, клеточные ансамбли, сообщающие о наличии дифференцируемых сигналов, широко распространены по обширным областям мозга и постоянно обнаруживают два типа ответов на два различных стимула. Анализ показывает, что информация, содержащаяся в усреднённых ответах каждого из этих двух локальных клеточных ансамблей, достаточна для различения двух периферических сигналов. Ансамбль из многочисленных реагирующих клеток обнаруживает поразительную стабильность в ответах, говорящую о существовании неизменного типа разрядов в ответ на каждый отдельный стимул. Напротив, активность одиночной клетки в высшей степени вариативна, проявляясь в виде разнообразных типов ответов при действии тех же самых стимулов. Эти факты говорят о том, что наборы разрядов обширных нейронных ансамблей служат гораздо более надёжным источником информации, чем наборы разрядов одиночных клеток. V обученных животных наблюдалось поразительное сходство в форме волн электрических ответов, регистрируемых в различных мозговых структурах. Анализ степени наклона кривых выявил, что эти сходные ответы широко распространены в головном мозгу, исключая проводящие пути. Динамическая природа этого феномена была проиллюстрирована фактами нарушения сходства при ошибочных ответах или при предъявлении новых стимулов. Эти факты свидетельствуют о том, что сходств» электрической активности в различных областях мозга отражает протекание некоторого эндогенного процесса, происходящего при научении, а не пассивно проявляющегося при действии физического стимула, как такового» (John and Morgades, 1969. — Такое понимание процесса подкрепления начинают признавать даже бихевиористы. Так, в недавнем обзоре (Perkins, 1968) утверждается, что концепция «привлекательности» объединяет целый ряд ранее разрозненных фактов. Согласно этой теории, «все классические условные рефлексы могут быть описаны как подготовительные ответы, которые повышают характерную привлекательность комплекса стимулов, совпадающих с предъявлением безусловных стимулов». Другие авторы употребляют термин «желательность», «побудительная ценность» или «побуждение»; я пользуюсь в этой книге термином «интерес» как субъективным эквивалентом «привлекательности». Однако общий смысл: этих терминов сводится к тому, что назначением процесса подкрепления является обращение к некоторым предшествующим состояниям, к некой компетентности организма, что в свою очередь приводит к изменению этих состояний. Компетентный мозгЧто же тогда представляют собой эти состояния компетентности, которые регулируют поведение? Это иерархически организованные механизмы (логические модули), включающие серво-процессы, программы или планы, направленные на достижение внешнего эффекта, действия. В главах XII и XIII была подробно описана нейронная организация механизма компетентности, участвующего в таких действиях, как, например, забивание гвоздя; анатомические структуры с однозначным соответствием между мышцами и корой мозга организуются в образ, который контролирует любые движения, диктуемые внешним или «силовым нолем» для достижения результата. Этот результат кодируется в образе, в состоянии, в микроструктуре медленных потенциалов соединений; всё это происходит посредством оценки мгновенных экстраполяции тех изменений в текущем процессе регулирования, которые появляются при сопоставлении соответствующих силовых полей с достигнутым результатом. Несколько более простой, но аналогичный механизм может быть описан по отношению к другим, более «внутренним» состояниям компетентности. Если у животных структуры ствола мозга (например, гипоталамус) стимулируются дискретными импульсами, возникают отдельные движения, имеющие отношение к еде, питью и так далее. Участки, ответственные за подобные отдельные движения,
Это предположение родилось в результате серии изобретательных экспериментов, выполненных Э. Валенстайном, которые, однако, в свою очередь поставили ряд новых вопросов. В этих экспериментах электроды вживлялись в ту область гипоталамуса крыс, электрическое раздражение которой вызывает движения еды, питья и попыток грызть кусок дерева. Когда один из таких поведенческих актов наблюдался особенно явно и стабильно, внешние условия эксперимента менялись таким образом, что первоначальный результат нельзя было получить. Так, в опыте создавались такие условия, когда в течение целой ночи животное через нерегулярные интервалы подвергалось стимуляции. На следующее утро та же стимуляция ранее раздражаемого участка вызывала либо первоначальное, либо изменённое поведение, что зависело от внешних условий, и, когда были испробованы все возможные варианты, обнаружилось, что часто доминировали новые выработанные движения. Валенстайн пишет:
Вторая серия опытов была проведена после применения различного числа тренировочных стимуляций в условиях отсутствия целевого объекта, к которому прежде стремилось животное… в большинстве случаев вторично стимулированное поведение появлялось так же закономерно, как и прежнее. Во время предъявления контрольного теста, когда одновременно присутствовали все три целевых объекта, в большинстве случаев с равной? вероятностью наблюдались два типа поведения. Как правило, один тип поведения возникал при действии первых трех—четырёх серий стимулов, а затем возникало другое поведение в приблизительно одинаковых по длительности сериях опытов. При предъявлении контрольных тестов животные обнаруживали несколько видов поведения за время предъявления некоторых из 20 стимулов. Мы хотели выяснить, станет ли доминирующим первый тип поведения, если мы дадим серию контрольных тестов. Хотя в этих опытах наблюдались разнообразные виды поведения, мы пришли к заключению, что как только закреплялся второй вид обусловленного стимулами поведения, печальное поведение, вызванное стимуляцией, переставало быть доминирующим. И, действительно, у ряда животных обнаружилось явное преобладание второго вида поведения во время предъявления контрольных тестов. (Valenstein, Cox and Kakolewski, 1969. — Валенстайн объяснил эти результаты тем, что стимуляция гипоталамуса не вызывает специфических мотивационных состояний типа голода или жажды. Исходя из фактов взаимодействия двух типов явлений — эффекта самораздражения по Олдсу и эффекта вызова с помощью стимуляции определённого вида поведения (еды, питья или попыток грызть дерево), — он приходит к выводу, сходному с тем, который был изложен в
Однако, предполагая, что стимуляция мозга не вызывает состояния жажды, Валенстайн может выбросить вместе с водой и ребёнка. У. Роберте (1969) указывает, что сильная электрическая стимуляция может разрушить чётко организованное поведение и расширить гипоталамическую зону, при раздражении которой постоянно наблюдаются частные движения, включённые в тот или иной поведенческий акт. Роберте обвиняет Валенстайна в том, что он стоит па позициях Лешли, отрицая специфичность нервных центров. Эта позиция, как он считает, должна быть пересмотрена, потому что, как отметил П. Милнер (Roberts, 1960. — Этот аргумент не нов, однако смысл его изменился. Почти сто лет поборники теории локализации функций спорили со сторонниками теории пластичности мозга; полем боя были зрительная и моторная кора. Теперь поле сражения переместилось в гипоталамус. Как при анализе восприятия и действия, решение вопроса
Общий вход, интеграция создаётся, конечно, в экспериментах Валенстайна благодаря комбинации электрической стимуляции моэга и подкрепления от окружающих условий, что — согласно концепции, изложенной в этой книге, — формирует окончательную структуру поведевия, которая опирается на динамическую систему медленных потенциалов соединений. Эти микроструктуры медленных потенциалов, эти кратковременные установочные состояния и регулируют поведение. Конечно, эти микроструктуры не обязательно локализованы в стволе мозга. Активирующий латеральный гипоталамический механизм является основным перекрёстком путей, проводящих сигналы от многих областей мозга, включая и кору больших полушарий. Результаты, полученные Валенстайном, как и повседневные наблюдения, говорящие о том, что заученные навыки сохраняются надолго, требуют, однако, более стабильной основы, чем эти кратковременные состояния, создаваемые микроструктурами соединений. Субстратом окончательного образа результата не могут быть чисто электрические процессы. Должны возникать Таким образом, мы вернулись к вопросам, рассмотренным во II главе. В следующей главе мы ещё раз обсудим эти вопросы и проанализируем модификацию поведения с точки зрения мотивации процессов памяти, то есть проблему подкрепления. РезюмеВ моторных механизмах мозга кодируются условия внешней среды, которые организуют поведение. Для решения проблемы, как это происходит, одинаково важны и работа нейрофизиологов, и усилия бихевиористов, направленные на изучение модификации поведения в результате научения, | |||||||||||||||||
Оглавление | |||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| |||||||||||||||||