Поиски энграммыОдной из важных особенностей состояний мозга является их пластичность. В модификации мозговых состояний выражается самое основное свойство нервной системы, её способность образовывать временные связи. Во второй главе будет более детально рассмотрена ткань мозга, в которой формируется микроструктура медленных потенциалов, с точки зрения её пластичности. До недавнего прошлого, несмотря на значительные усилия исследователей, ещё не было прямых доказательств нейронной модификации, возникающей в результате индивидуального опыта организма. В 1950 году Лешли закончил свою известную работу, посвящённую исследованию энграммы, следующими словами: «Анализируя данные, касающиеся локализации следов памяти, я испытываю иногда необходимость сделать вывод, что научение вообще невозможно. Тем не менее, несмотря на такой довод против него, научение иногда происходит» (Lashley, 1950. — Сейчас положение изменилось. Интенсивное изучение того, каким образом изменяется мозг под влиянием индивидуального опыта, привело к открытию нескольких направлений в исследовании проблемы. Мы рассмотрим данную проблему в три этапа. Настоящая глава будет посвящена развитию долговременных изменений в нервной системе в процессе научения. В Почему проблема биологии памяти, поиски доказательств того, что опыт вызывает постоянные изменения в нервной ткани, оказались столь трудными? Когда в каких-либо органах тела возникают со временем нормальные структурные изменения, мы говорим о росте и развитии ткани. Но в мозгу число нейронов не увеличивается после начального периода, непосредственно следующего за рождением. Таким образом, практически все развитие поведения и научение происходит при отсутствии каких-либо указаний на рост и развитие в нервной системе (см. Pис.
Этот парадокс может быть разрешён тремя направлениями исследований. Первое отрицает важность того факта, что нейроны не способны к воспроизведению, и делает попытку установить, что существует Рост нервной ткани
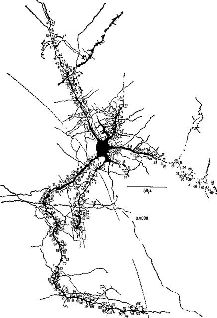
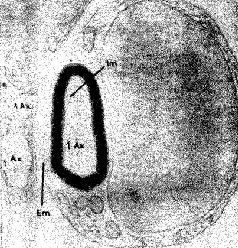
Возможность роста нервной ткани не отрицается полностью тем фактом, что нейроны не способны к воспроизведению, как другие клетки тела. Нейроны отличаются друг от друга своими длинными волокнами, протяжённостью разветвлений на теле клетки и теми амебовидными структурами на концах, названными конусами роста, которые можно наблюдать в культуре ткани и даже in situ (см. Pис. Конус роста толкает и давит на находящуюся перед ним ткань и может открывать путь, по которому он затем сможет медленно течь, удлиняя конец нервного волокна. Таким образом, рост может происходить при условии наличия свободного пространства, когда же его нет, амебовидный конец сокращается только для того, чтобы снова и снова продвигаться вперёд. В центральной нервной системе рост обычно исключён потому, что элементы ткани расположены очень близко друг к другу. Правда, возникли большие сомнения в этом, так как результаты электронной микроскопии показали, что внеклеточное пространство, которое было найдено во всех частях организма, существует и вокруг нейронов. Поэтому исследователи мозга стали наблюдать за ростом нервного волокна, создавая в мозгу свободное пространство. До недавнего времени такие попытки не приводили к значительным результатам: обычная реакция мозговой ткани на инсульт состоит в разрушении повреждённых частей и разжижении их, в образовании полости или кисты, оболочки которых препятствуют любой попытке проникновения в них. Однако современная техника дала исследователям инструмент, позволяющий избежать этой обычной реакции. Циклотрон излучает радиацию, которая резко замедляет процессы, происходящие в мягкой ткани. Таким образом, описанные разрушения можно произвести локально и только на том участке, на который направляется разрушающее воздействие. С помощью этого метода можно разрушать определённые слои коры взрослого кролика без видимого повреждения соседних. При этом методе повреждение ткани достаточно локально, так что обычно наблюдаемая реакция на большие разрушения нервной ткани не имеет места. Таким образом, создаётся пространство для активного роста нервных волокон (см. Pис.
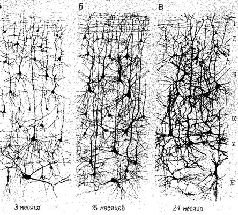
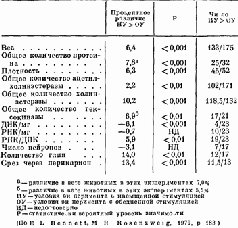
Срезы, сделанные спустя несколько недель или месяцев после такого повреждения, показывают, что сохранившиеся волокна становятся толще (Kruger, 1965). Это утолщение, Эти эксперименты, конечно, не могут показать, является ли рост действительно результатом жизненного опыта; для этого должен быть проведён другой тип исследований. Чтобы проверить это, условия содержания крыс менялись таким образом, что одни из животных получали много стимулирующих воздействий (игра, решение задач), тогда как другие подвергались сравнительно ограниченной стимуляции. Иногда это ограничение касалось либо одной, либо другой сенсорной модальности (например, крысы содержались в полной темноте), чтобы проверить возможность различного воздействия на некоторые области мозга. Сравнение мозга животных, содержащихся в различных условиях, показало, что стимуляция вызывает заметное утолщение соответствующих отделов коры у животных, жизненный опыт которых был богаче (Krech, Rosenzweig, Bennett and Diamond, 1964). Тщательный гистологический анализ утолщенной коры показал — как и следовало ожидать, так как число нейронов не увеличивается после рождения, — что количество нервных клеток на единицу объёма, по существу, несколько уменьшилось. Однако было отмечено увеличение числа разветвлений базальных дендритов, увеличение числа и расширение дендритных шипиков — маленьких волосковых выступов, которые, как предполагают, являются местом активного соединения нейронов, распространяющих свои волокна горизонтально по коре. Большая же часть утолщения происходит за счёт увеличения не нервных клеток — глии (см. Pис.
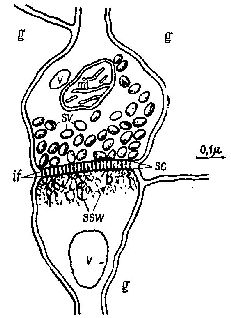
Роль нейроглииВторым центром внимания при изучении проблем сохранения следов памяти стали имеющиеся в нервной ткани не нервные клетки, названные нейроглией (или просто глией). Глиальные клетки происходят из той же эмбриональной ткани, что и нервные. Длительное время считалось, что назначение глии состоит исключительно в том, чтобы служить источником питания для нейронов. Эта функция глии была достаточно хорошо доказана (например, Hyden, 1965). Глия идеально расположена для выполнения этой роли — некоторые из её клеток (астроциты) окружают кровеносные сосуды, и таким образом посредством кровообращения может происходить метаболический обмен. Другие (олигодендроглия), имеющие небольшое количество ветвей, плотно оплетают нервные волокна, особенно на концах, заполняя таким образом межклеточное пространство вокруг нейронов на этих участках. Кроме метаболической функции, олигодендроглия способствует образованию изолирующего материала (миелина), который покрывает крупные нервные волокна (Рис.,
Другое важное назначение олигодендроглии состоит, Эти специфические репаративные шванновские клетки в эмбриогенезе происходят из того же источника, что и нервная и глиальная ткани. Более того, эти шванновские клетки обвиваются вокруг нервных волокон (см. Pис.
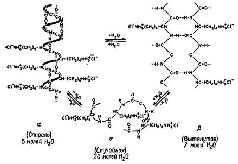
Эти данные и привели большинство исследователей к объединению олигодендроглии и шванновских клеток в один класс. Вполне возможно, следовательно, что олигодендроглия направляет рост нейронов в центральной нервной системе, подобно тому как это делают шванновские клетки в периферической нервной системе. Тот факт, что глия обвивается вокруг окончаний нервных волокон, ставит перед нами интересную проблему. За исключением тех случаев, когда соединение между нервами осуществляется посредством особых структур, называемых синапсами, изменения электрических потенциалов, возникающие в нейронах, вызывают потенциалы Насколько же сильно влияние этих клеток на конфигурации, образуемые потенциалами соединений? Хотя на такие вопросы ещё не получено ответа, они заставляют предположить, что происходящие в глии изменения могут постоянно менять активность групп нервных клеток, с которыми она так тесно связана. Химическая пластичностьРяд экспериментов прямо указывает на участие глии в механизме памяти. Одним из веществ, обеспечивающим взаимодействие между глией и нейронами, оказалась рибонуклеиновая кислота (РНК — химическое вещество, определяющее конфигурацию-протеинов), которая сама является производной дезоксирибонуклеиновой кислоты (ДНК), молекулы генетической памяти. Фактически эти исследования показали, что при стимуляции нейронов вырабатывается больше РНК, чем в любой другой ткани тела (Hyden, 1961). После прекращения стимуляции и уменьшения выработки РНК в нейроне большие концентрации молекулярно сходной РНК начинают появляться в находящейся по соседству глии (Hyden, 1969). Эти эксперименты, проведённые на мозге кроликов и крыс, были усовершенствованы введением выполняемой под микроскопом процедуры отделения глии от нейронов в вестибулярном ядре — группе нейронов, контролирующей поддержание равновесия животного. Стимуляция этого механизма «равновесия» первоначально осуществлялась следующим образом: животное помещалось в центрифугу и вращалось в ней. В более поздних экспериментах от крыс требовалось, чтобы они карабкались по наклонно натянутой проволоке. При успехе они достигали платформы с едой, при неудаче падали на расположенную внизу клетки решётку, по которой пропускался слабый электрический ток. На различных стадиях эксперимента микрохимическими методами определялась концентрация РНК в вестибулярных нейронах и глии животных. Во время пассивного вращения в центрифуге общее количество РНК в нейронах увеличивалось; увеличение содержания РНК было обнаружено
По-видимому, никто не сомневается в том, что при физиологической стимуляции нервов или во время выполнения Этот взгляд на функцию РНК предполагает, что сохранение следа происходит в других макромолекулах, таких, как полипептиды, протеины, липопротеины и мукоиды (см. Bogoch, 1968; Glassman, 1967). Все эти молекулы имеют сложное строение и синтезируются в результате многих метаболических реакций. Следовательно, можно провести эксперименты, в которых тормозящие обмен вещества действовали бы на то или другое звено цепи синтеза. Эти вещества можно вводить как до
С помощью структурных изменений можно хорошо объяснить временную память, связанную с такими образами, которые характерны для лиц с «фотографической» (или «эйдетической») памятью. Один такой эйдетически одарённый студент недавно был обнаружен в Гарварде. В результате экспериментальной проверки было показано, что он способен сохранять в памяти каждую деталь своих зрительных восприятий в течение 8 дней. Вторая часть книги будет в значительной мере посвящена рассмотрению этой временной спирализации молекул. Изучение структурных изменений мозговой ткани сейчас едва ли возможно, однако техника развивается такими темпами, что эта область исследования многое обещает уже в ближайшем будущем (Sjostrand, 1969). Некоторые исследователи продолжают придерживаться гипотезы, что РНК является непосредственным хранилищем памяти, несмотря на серьёзные доводы против этой гипотезы. Они основывают свою точку зрения яа весьма спорных экспериментах с «переносом», которые были проведены на планариях, крысах и обезьянах. В этих экспериментах экстракт РНК от контрольных животных и животных, обучавшихся выполнению задания, вводили необученным животным. Те из них, которым вводили РНК от животных, имевших опыт обучения, иногда, но не всегда решали задачи быстрее по сравнению с теми, которым вводили РНК от контрольных животных. Согласно некоторым данным, эффект различного действия РНК «обученного» и «контрольного» животного исчезал в результате обработки экстракта РНК веществом, избирательно разрушающим РНК (см. обзор McGonnell, 1970). Сторонники химической гипотезы сохранения следов, оказавшись перед необходимостью выбора между двумя возможными решениями, в конце концов остановились на нейронных медиаторах. Уменьшение скорости и амплитуды нервных импульсов, когда они достигают окончаний аксонов, возникает
Память могла бы зависеть от лёгкости, с которой высвобождается медиатор, или от его количества. Таким образом, можно провести такие эксперименты — и они действительно проводятся, — когда медиатор нейтрализуется в результате действия фармакологических веществ или его разрушение блокируется во время тренировки животного (Deutsch, Hamburg and Dahl, 1966). В результате таких действий у крыс нарушается процесс научения, но, как Индукция как модель хранения следов памятиМоя реакция на изобилие экспериментальных данных, полученных при исследовании энграммы в 1960 году, двойственна. Во-вторых, простая модель следов, предполагающая постоянную модификацию мозговой ткани на нейронном уровне, может быть сформулирована уже сейчас. Такая модель должна обеспечить не только возможность сохранения следов, но и доступность считывания зафиксированного изменения. Наконец, такая модель должна быть основана на уже имеющихся данных и отвечать здравому биологическому смыслу. В настоящее время наиболее вероятным источником такой модели может служить аналогия с процессом, происходящим во время эмбрионального развития организма (см. Pис.
Такие структуры, как глаз, формируются только тогда, когда ткань, из которой они развиваются, соответствующим образом стимулируется. Большая часть эмбриональной ткани эквипотенциальна, то есть ДНК во всех клетках тела, по существу, одна и та же. Это означает, что потенциал первоначально подавлен, находится в состоянии «репрессии». «Дерепрессия», или освобождение потенциальных возможностей, происходит тогда, когда появляются соответствующие условия. Определение эмбриологами того, каковы эти условия, заняло почти целое столетие. В настоящее время хорошо известно, что РНК и некоторые эндокринные секреты являются индукторами такой «дерепрессии». Маловероятно, что эти химические соединения несут в себе всю информацию, необходимую для индукции; большая доля специфики процесса должна быть отнесена за счёт субстрата. Например, РНК, выделенная из печени телёнка, может быть использована как индуктор для хрусталика глаза крысы. Отношения между индуктором и субстратом,
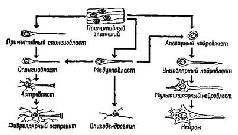
Данные о роли РНК в сохранении следов памяти в настоящее лремя могут быть объяснены при помощи модели, основанной на этом эмбриональном процессе индукции. Модель предполагает, что возбуждение нервных структур сопровождается образованием РНК. Эта нейронная РНК вызывает изменения в окружающей «лигодендроглии, в результате чего начинается химический процесс между нейроном и глией, характеризующийся реципрокными отношениями изменений нейронной и глиальной концентрации РНК (и множества метаболитов). Происходит изменение в функциональном взаимодействии между глией и нейроном. На первом этапе может возникнуть соответствующее изменение концентрации РНК в глиальной клетке, которое затем в течение длительного времени будет вызывать изменение в структурах липидов, протеинов и липопротеинов во всех больших молекулах, составляющих мембраны, через которые осуществляется взаимодействие нейрона с глией.
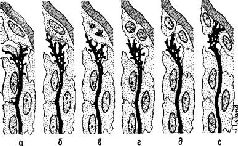
Такие макромолекулярные изменения могут влиять на лёгкость освобождения и разрушения нейронных медиаторов. Эти структурные изменения обратимы и могут стираться или быть вытеснены другими изменениями. Однако, если при неоднократном повторении одного и того же типа воздействия’ изменения в молекулярной структуре будут продолжаться достаточно долго, они вызовут эффективные изменения мембранной проницаемости, которые в свою очередь способствуют появлению в Возбуждённом нейроне большого количества РНК, метаболитов и медиаторов, оказывающих влияние на окружающую его глию в том месте, где происходит деление глиальной клетки. Конус роста нейрона, освободившийся от инкапсулированной глии, свободно воздействует на недавно сформировавшиеся дочерние клетки в образует новые связи с нейронами. Таким образом, клеточное деление олигодендроглии направляет конусы роста нейронов центральной нервной системы во многом так же, как на периферии их рост направляют соответствующие шванновские клетки (см. Pис. Консолидация следовЛюбое изменение глиально-нейронной организации следует рассматривать как изменение в микроструктуре активности медленных потенциалов соединений. Как мы уже говорили, такие изменения могут быть временными либо вести к возникновению более устойчивых изменений в нервной системе, связанных с ростом нейронов. Имеется много данных, свидетельствующих о том, что для фиксации в мозгу следов памяти — энграмм — требуется время. Эти данные вполне согласуются с индукционной моделью хранения следов памяти, потому что они указывают на возможность выделения по крайней мере двух — если не больше — процессов в зависимости от времени их появления. После достаточно сильной травмы головы человек не способен воспроизвести в памяти события, происходившие в период, непосредственно предшествовавший травме. Длительность этого периода зависит от тяжести травмы. Этот феномен, названный ретроградной амнезией, может быть использован для изучения фиксации следов, то есть для изучения процессов консолидации. Самым распространённым методом, чтобы вызвать ретроградную амнезию в лабораторных условиях, является применение электроконвульсивного шока на крысах, хотя используются также и другие методы, такие, как удар по голове, быстрая анестезия с помощью эфира или введение определённых веществ, вызывающих амнезию. Полученные данные говорят о том, что, чем скорее после того, как возник навык, вызываются конвульсии, тем сильнее их тормозное, амнестическое действие на последние из навыков, приобретённые в этом эксперименте. Так, в одном исследовании (Chorover and Schiller, 1965) максимальный эффект отмечался в тех случаях, когда конвульсии следовали не позднее, чем через 15 секунд после условной реакции избегания. Эффект практически исчезал, когда проверку выработки условного рефлекса и конвульсии разделял интервал длительностью 1 час. Эффект в значительной мере зависит от сложности задания, которое нужно было выполнить. Однако это ещё не всё. При соответствующих условиях возникает более сложный, двухэтапный процесс консолидации. Допустим, испытуемый должен нажимать на рычаг, чтобы получить в качестве вознаграждения пищу, предъявляемую со слегка меняющимися интервалами. В Результаты этих экспериментов по консолидации следов памяти можно интерпретировать, в соответствии с индукционной моделью, следующим образом: сразу же после испытания появление шоковых конвульсий, анестезия и так далее препятствуют метаболическому обмену, происходящему между нейроном и олигодендроглией, тормозя индукцию, вызываемую путём секреции РНК в нерве и сопровождающуюся её активацией в глии. Следовательно, образование энграммы прекращается и припоминание становится невозможным. Напротив, когда конвульсии возникают позднее, связанная с РНК фаза процесса индукции оказывается уже завершённой. В этом случае временно приостанавливается деление глиальной клетки, амебовидные движения окончаний нейронов временно прекращаются и конус роста втягивается назад. Однако при восстановлении этих механизмов процесс запечатления энграммы продолжается с того момента, когда он был прерван, и при завершении этого процесса становится возможным припоминание. Чтобы твёрдо установить, что консолидация следа памяти действительно происходит, необходимо найти такой метод, который улучшал бы научение. Это, в сущности, нетрудно. Когда крысам вводили сульфат стрихнина или аналогичные возбуждающие вещества не ранее чем за 10 минут до начала или 30 секунд спустя после пробы на прохождение лабиринта или на выработку дифференцировки, то навык у них вырабатывался быстрее, нем у контрольных; консолидация, таким образом, происходила скорее (McGaugh and Petrinovich, 1959). Эти эксперименты служат красноречивым свидетельством того, что консолидация является необходимым условием возникновения следа памяти. Консолидация возникает в мозгу, но что же именно происходит во время этого процесса, который делает возможным запоминание? Ответ на этот вопрос могут дать следующие наблюдения. В моей лаборатории на различные области коры мозга обезьян наносилась алюминиевая паста для создания локальных раздражений, которые в отдельных случаях вызывали эпилепсию. Эти раздражения проявлялись в изменённой электрической активности — в возникновении патологических медленных волн и спайковых разрядов. Хотя такие нарушения не мешали обезьянам помнить решение задач, с которыми они неоднократно имели дело до раздражения, эти нарушения замедляли первоначальный процесс выработки данного навыка почти в пять раз (Pribram, 1966). Более того, способность вырабатывать такие навыки в целом не подвергалась нарушению; оно касалось только тех задач, выполнение которых было связано с определённой частью мозга, и ограничивалось лишь начальным периодом процесса выработки навыков, предшествующим появлению первых признаков научения. Таким образом, вызываемые в этом опыте раздражения не всегда блокируют консолидацию следов. Может быть, одиночная энграмма, ограниченная лишь одним нейронным фокусом, является недостаточно эффективной, чтобы проявить себя? Большинство результатов экспериментов по научению у человека показывает, что, для того чтобы запомнить задание, необходимо его многократное повторение (Trabasso and Bower, 1968). Во время повторения происходит такое распределение повторяемого материала, что он включается в целую систему связей предварительно зафиксированного опыта. Таким образом, результаты экспериментов с нарушениями, вызванными раздражением, можно объяснить тем, что процесс повторного возникновения и распределения энграммы был задержан. Проверку этой интерпретации можно осуществить путём сравнения результатов научения обезьян с повреждениями мозга в условиях распределённого концентрированного научения. Гистологический анализ мозговой ткани, подвергшейся воздействию алюминиевой пасты, показывает наличие спутанных клубочков нервных волокон, во многом напоминающих соответствующие образования в периферических нервах, когда их рост не направляется соответствующей колонкой шванновских клеток. Возможно ли, чтобы это воздействие избирательно уничтожало олигодендроглию, вызывая ненаправленный рост нервных волокон? Химический анализ ткани, в которую была введена алюминиевая паста, конечно, не осуществим. Однако, чтобы получить ответ на этот вопрос, был проведён остроумный эксперимент (Morrell, 1961,
Разрушение, вызванное раздражением в одном полушарии головного мозга, через несколько месяцев вызывает в контралатеральной коре появление «зеркального очага» изменённой электрической активности, который возникает при участии мозолистого тела. Этот «зеркальный очаг» непосредственно не испытывал разрушающего химического воздействия, но обладал всеми признаками, характерными для первичного очага разрушения, вызванного раздражением. Существенно ли меняется РНК в «зеркальном очаге» по сравнению с нормальной тканью мозга? Может ли изменённая РНК быть непосредственной причиной этого раздражения и последующих трудностей в восстановлении следа памяти? Индукция в памяти, так же как и индукция в эмбриогенезе, является, видимо, многоэтапным процессом, который требует времени для своего завершения. Можно ожидать, что каждая стадия в таком процессе имеет своё уязвимое место, которое можно выявить с помощью соответствующего метода, применённого в критический момент. Уже многое известно о «критических периодах» в развитии эмбрионов и поведения в первые годы жизни. Определение природы критических, то есть сензитивных, периодов в процессе образования следов памяти представляет собой богатую область для исследования и экспериментов. Можно ещё многое сказать — и это будет сделано — о консолидации и модели индукции, но сначала нам нужно составить более полное представление о принципах работы мозга РезюмеДлительно сохраняющиеся изменения происходят в соединительных аппаратах мозговой ткани. Хотя зрелые нервные клетки не делятся, механизм постоянной модификации мозговой ткани обладает многими свойствами механизма дифференциации эмбриональной ткани. В обоих случаях можно наблюдать экспериментально вызванный направленный рост новых нервных волокон, который меняет пространственную структуру связей между нейронами. Следовательно, долговременная память является скорее функцией соединительных структур, чем функцией процессов в самой нервной клетке, генерирующей нервные импульсы. | |||||||||||||||||||||||||||||||||
Оглавление | |||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| |||||||||||||||||||||||||||||||||