Подкрепление как индукцияСформулированное в главе XIV утверждение о том, что процесс подкрепления индуцирует изменения в механизмах компетентности мозга, нуждается в тщательной проверке. Имеется ли какое-либо сходство между процессами индукции, изучаемыми эмбриологами, и процессами подкрепления, изучаемыми бихевиористами? В главе II процесс индукции рассматривался в качестве-модели хранения следов памяти. Какие нервные механизмы могли бы приходить в действие во время процесса подкрепления, который вызвал бы процесс индукции в мозгу взрослого организма? Наиболее спорная трактовка подкрепления, предлагавшаяся как бихевиористами, так и психоаналитиками, заключалась в попытке рассматривать подкрепление как фактор, уменьшающий влечение. Эта концепция основывается на двухфакторной теориш влечения: физиологические потребности приводят к состоянию напряжения в организме, которое проявляется в повышении общей» активности или в усилении нервной активации; поведение, направленное на уменьшение этого напряжения, и есть подкрепление. Некоторые авторы (например, Sheffield et al., 1955) считают, что организмы, напротив, стремятся к повышению напряжения и что изменения поведения сопровождаются ростом: напряжения. Этот аргумент устраняет необходимость во втором факторе. Однако влечение и подкрепление по-прежнему рассматриваются как взаимосвязанные явления. Между тем Эстес (1958) привёл убедительные доводы в пользу концепции, согласно которой поведение строится по схеме «побуждение — стимул», а не по схеме «побуждение — напряжение», однако эта концепция: оставляет без ответа как раз вопрос о том, какие же побуждающие стимулы управляют поведением. Ответ на этот вопрос сходен с тем, который был дан в предыдущей главе, а именно вопрос, что такое «стимул», — не так прост, как кажется. Побуждающий стимул, как и сенсорный стимул, обусловлен работой корригирующего сервомеханизма, или гомеостата. Гомеостаты снабжены рецепторами, чувствительными к возбуждению в системе «внешний мир — внутренняя среда». Особые зоны мозга, чувствительные к осмотическим колебаниям, изменениям температуры, содержанию эстрогена, глюкозы и уровню углекислоты, находятся вокруг желудочков мозга; они расположены по средней линии; зти области связаны с механизмами, контролирующими поступление и расход агентов, к которым они чувствительны (см. Главу IX, X). Помимо этих центральных механизмов, в гомеостатических процессах принимают участие также другие, более периферические механизмы. Центральные гомеостатические процессы часто дополняются вторичными механизмами, которые обеспечивают более точно дозированную регуляцию указанных агентов. Примерами могут служить сокращения желудка в механизме пищевой регуляции во время голода или сухость во рту при жажде, а также регуляция циркуляции крови в сосудах кончиков пальцев для большего или меньшего охлаждения тела. Основные гипоталамические терморегуляторы дифференцированно управляют температурой пальцев посредством кровообращения. Если взять в качестве модели голод, то можно видеть, что мозговой гомеостатический механизм с его центральными и периферическими чувствительными аппаратами имеет два реципрокно действующих компонента. Один из них сигнализирует о недостатке какого-либо агента и запускает регуляторный процесс, другой сигнализирует о насыщении организма и останавливает этот процесс. Можно также видеть, что фаза запуска регуляторного процесса характеризуется наличием потребности, а фаза прекращения вызывает состояние аффекта. Следовательно, оказывается, что потребности и аффекты, чувство заинтересованности представляют собой мотивационные и эмоциональные (в отличие от перцептивных) стимулы, иначе говоря, то, что бихевиористы называют влечениями. На языке этой книги потребности и аффекты являются управляющими образами, индикаторами процессов, связанных с состояниями мозга и таким путём: влияющих на организацию поведения во времени (см. Главу II). Каким образом механизмы, обеспечивающие состояния интереса, потребности и аффекта, модифицируют поведение организма и служат подкреплением? Путём вовлечения механизма памяти, которым обладает организм. В главе II была сформулирована гипотеза, согласно которой структуры, ответственные эа долговременную память, как бы стимулируются в мозгу, подобно тому как стимулируются тканевые структуры в процессе эмбрионального развития. Внешнее сходство между процессами индукции, изучаемыми в эмбриональных тканях, и процессами подкрепления, изучаемыми в ситуации выработки условных рефлексов, легко сформулировать в следующих положениях: - Индукторы пробуждают и организуют генетический потенциал организма. Подкрепляющие агенты пробуждают и организуют поведенческие возможности организма. Б) Индукторы сравнительно специфичны в отношении действия, которое они вызывают, но, как правило, неспецифичны по отношению к индивидам и тканям. Подкрепляющие агенты довольно специфичны по отношению к тому поведению, которое они обусловливают, но, как правило, неспецифичны по отношению к индивидам и задачам.
- Индукторы детерминируют широкий набор индуцированных эффектов, детали уточняются процессами, протекающими в субстрате, по мере развития ткани. Подкрепляющие агенты определяют решение проблемной ситуации, детали поведенческого репертуара (конкретные формы поведения), используемого для достижения результата, сугубо индивидуальны.
- Индукторы не являются просто агентами, запускающими развитие, они представляют собой нечто большее, чем мимолётные стимулы. Подкрепляющие агенты не просто агенты, запускающие поведение; они представляют собой особый класс стимулов.
- Индукторы должны контактировать с субстратом, для того чтобы быть эффективными. Совпадение во времени — обязательное требование для того, чтобы реализовать подкрепление.
- Простой контакт, хотя он и необходим, недостаточен, чтобы вызвать эффект индукции. Индуцируемая ткань должна быть готовой к реакции, должна быть компетентной. Простое сочетание, хотя оно и необходимо, недостаточно, чтобы вызвать подкрепление; подготовленное состояние, недостаток информации, депривация, готовность, контекст, ожидание, внимание, создание гипотезы — вот только некоторые из терминов, используемых для описания тех факторов, которые создают компетентность организма, без чего подкрепление не может стать эффективным.
- Индукция обычно осуществляется благодаря двустороннему взаимодействию — благодаря химическому взаимодействию. Подкрепление является наиболее эффективным в оперантной ситуации, где последовательность действий самого организма немедленно используется как регулятор его последующего поведения.
Нейрохимия подкрепления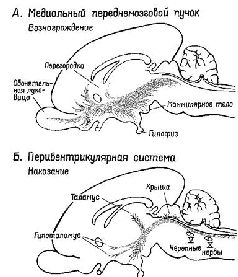 | | Рис. XV–1 Схема медиального переднемозгового пучка (верхний рисунок — система запуска) и перивентрикулярной системы волокон (нижний рисунок — система остановки) в мозгу млекопитающих (сагиттальный разрез) (Stein, 1968, Le Gros Clark et al, 1938) | 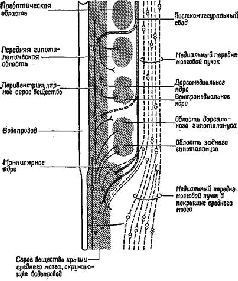 | | Рис. XV–2. Схема медиального переднемозгового пучка (пунктир) и перивентрикулярной системы волокон в горизонтальной плоскости. На схеме гипоталамус представлен как три продольно расположенных столба — перивентрикулярный поясной слой, медиальная зона и латеральная зона. Пучки волокон в перивентрикулярной и латеральной зонах примыкают к медиальной зоне, которая лишена крупных пучков волокон, но содержит несколько известных ядер. Наклонные линии обозначают систему свода (Stein, 1968; Sutin, 1966). |
После всего, что уже сказано, остаётся вопрос, указывает ли описанное выше сходство процессов индукции и подкрепления на сходство их механизмов? С моей точки зрения, да. Какие же факты это подтверждают? Какие нервные процессы начинают действовать во время индукции? Для этого требуются анатомические пути, функционирующие во многих областях мозга, чтобы запустить в действие подкрепляющий «индуктор», химические агенты, стимулирующие реципиентную ткань к дифференциации. Пример подобного любопытного синтеза мы находим в нейрохимическом исследовании, проведённом Л. Стайном (см. Pис. XV–1, XV–2). «Все приведённые факты отвечают представлению о том, что механизм подкрепления содержит систему адренэргических Синапсов, которые в высшей степени чувствительны к фармакологическим воздействиям Усиление порадренэргической передачи в этих сииаптических аппаратах вызывает эффекты облегчения в поведении, ухудшение норадренэргической передачи подавляет поведение При всех условиях эти синаптические аппараты являются наиболее важным участком работы мозга, где амфетамин и хлорпромазин оказывают влияние на целенаправленное поведение.
Где локализуются эти синапсы? Сели в медиальном переднемозговом пучке действительно находится система вознаграждения, как указывалось выше, то данные адренэргические синапсы, очевидно, уже описаны группой сотрудников Каролинского института (Fuxe, 1965, Hillarp, Vuxe and Dahlstrom, 1966) Используя гистохимическую технику для обнаружения катехоламинов на клеточном уровне, эти исследователи сообщают о системе нейронов, содержащих порадреналин; клеточные тела этих нейронов берут начало в мезенцефальных отделах мозга, а волокна их восходит к медиальному иереднемозгопому пучку и оканчиваются в адренэргических синапсах гипоталамуса, лимбической доли и неокортекса… Используя совершенно иную технику, независимо от предыдущих авторов Геллер, Сайдеи и Мур (1966) подтвердили существование этой восходящей системы волокон. Эти исследователи разрушали медиальный переднемозговой пучок па одной стороне на уровне латерального гипоталамуса и затем проверяли изменение содержания норадреналина в различных областях мозга. Поскольку в медиальном переднемозговом пучке аксоны в большинстве случаев не перекрещиваются, оказалось возможным сравнить уровень содержания норадреналина на поражённой и интактной сторонах. Было обнаружено, что на поражённой стороне — и только в структурах пучка мозга — норадреналина было значительно меньше; контрольные разрушения медиального гипоталамуса не привели к существенному уменьшению норадреналина. Эти результаты подтверждают существование в медиальном переднемозговом пучке волокон, содержащих норадреналин; кроме того, авторы приходят к выводу, что эти волокна входят в восходящую систему, так как уменьшение уровня норадреналина наблюдается только в структурах, расположенных выше уровня поражения…» (Stein, 1968. — p. 110) 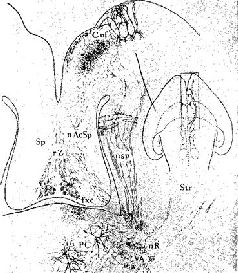 | | Рис. XV–3. Ростральные проекционные пути неспецифической талампческой системы, идущие через нижние ножки таламуса (часть медиального переднемозгового пучка) в кору. Аксоны от передних неспецифических полей, включающих парацентральное ядро (PC) и, возможно, медиальную часть вентрального переднего ядра (VA), проходят рострально через нижние ножки таламуса как неспецифические проекционные пути (nsp) и через медиальный сектор каудального отдела полосатого тела (Str) к основанию орбитофронтальной коры (Cof). Эти аксоны широко ветвятся в субгризеальном белом веществе, и некоторые из них лродолжаются в ядрах ограды [n] accumbens (nAcSp). Остальные сокращения означают: ретикулярное ядро таламуса (nR), столбы свода (fxc), ограда (Sp) и радиация Цукеркэндла (rZ). Эпидермальная нейроглия [g] тянется вдоль боковых желудочков. Горизонтально-наклонное сечение, модифицированная методика быстрого окрашивания по Гольджи. 50-дневный, частично демиелинизированный мозг мыши X 150. В репродукции масштаб сокращён до Х0.75 (Scheibel, 1967). | 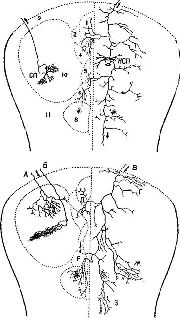 | | Рис. XV–4. Сравнение организации аксонных элементов специфической и неспецифической таламических систем. Верхний рисунок — таламофугальные элементы. Нейрон специфического вентробазального комплекса ядер (СП) посылает вертикально направленный без коллатералей аксон (10) к коре, исключение составляет лишь одна маленькая возвратная ветвь. Нейрон неспецифической системы (НСП) посылает аксон, который разветвляется на ростральные и каудальные отростки, богато снабжённые коллатералями и идущие ипсилатерально и контр-латерально. Нижний рисунок — нисходящие таламопетальные элементы. А и Б — аксоны, идущие от корковых нейронов к комплексу различных специфических вентральных ядер. А — характерная объёмная пограничная структура; Б — плоский древовидный нейрон, имеющий форму диска; В — аксон, идущий из коры к неспецифическим структурам. В диффузную, богатую коллатералями систему входят ветви, идущие к области ретикулярного ядра и к вентральному переднему ядру [Г], к неспецифическим структурам другого полушария (Д и Е), к заднему комплексу ядер [Ж] и мезодиенцефальным образованиям [3]. Цифры обозначают: 1 — parataential; 2-anterior ventral; 3 — interanteromedial; 4 — ranterior medial; 5 — paracentral; 6 — central lateral; 7 — central medial; 8 — centre median-parafascicularis complex; 9 — nreticularis. Рисунок составлен на основании изучения ряда срезов мозга крыс и мышей до методу Гольджи (Scheibel, Scheibel, 1967). | 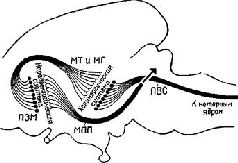 | Рис. XV–5. Схема, показывающая возможные взаимосвязи между механизмами вознаграждения и наказания, составленная на основе экспериментов Маргулиса, Стайна и другие (1967, 1968) по химической стимуляции. Стимулы вознаграждения «запускают» поведение, освобождая его от тормозящих механизмов с помощью перивентрикулярной системы (ПВС), причём имеет место следующая последовательность явлений:- Активация медиального переднемозгового пучка (МПП) с помощью стимулов, предварительно ассоциированных с наградой (или избежанием наказания) приводит к высвобождению норадреналина в амигдале и других подавляющих зонах переднего мозга (ПЗМ).
- Тормозное действие норадреналина подавляет активность, сокращая тем самым опосредованное холинэргином возбуждение медиального таламуса и гипоталамуса (МТ и МГ).
- Ослабление холинэргической синаптической передачи в МТ и МГ уменьшает активность перивентикулярной системы и таким образом снижает её тормозное влияние на моторные ядра ствола мозга (Stein, 1968).
| 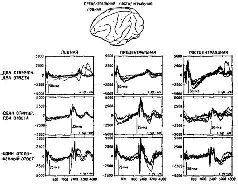 | | Рис. XV–6. Сравнение транскортикальной негативной вариации (TNV) — в лобной, прецентральной и постцентральной областях коры обезьяны резус во время выполнения различных заданий на ожидание. В ситуации предъявления двух сигнальных стимулов, требующих двух ответов, обезьяна должна нажимать на рычаг в пределах 400 мсек после включения светового стимула, держать рычаг опущенным в течение двух серунд и отпускать его в пределах 400 мсек после включения звука. В ситуации предъявления одного сигнального стимула и двух ответов обезьяна должна была спонтанно начинать опускание рычага без стимула, держать рычаг опущенным в течение двух секунд и быстро отпускать его при включении звука. В ситуации отсроченных реакций свет и звук предъявлялись с интервалом в 2 секунды; обезьяна должна была воздерживаться от ответа до тех пор, пока не появится звук. Каждая запись — усреднённые компьютером 40–50 ответов. В качестве подкрепления обезьяны получали за каждый правильный ответ маленькие кусочки пищи. Ср. с pис. XV–7 (Donchin et al., 1971). | 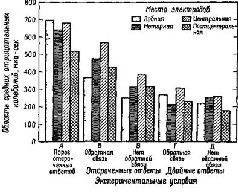 | | Рис. XV–7. Сравнение контингентных негативных отклонений (CNV) в различных участках мозга у человека в 5 различных экспериментах на время реакции. Во всех опытах предъявлялись 2 зрительных стимула с интервалом в 1,5 секунды. В экспериментах А — В испытуемый должен был нажимать на кнопку большим пальцем ведущей руки в ответ иа второй стимул. В экспериментах Г и Д от испытуемого требовалось нажимать на кнопку сразу же после включения первого стимула, держать её нажатой в течение интервала между стимулами и отпускать при появлении второго стимула. В качестве обратной связи использовался красный или зелёный свет, который зажигался после каждого правильного или ошибочного ответа. В экспериментах В — Д испытуемые должны были реагировать в пределах 350 мсек. В этих условиях ограниченного времени время реакции сокращалось настолько, что испытуемые допускали до 25 процентов ошибок. Область отрицательных колебаний оценивалась методом суммирования всех отрицательных отклонений (относительно исходной средней линии), возникающих в течение двухсекундного интервала, следующего за включением первого стимула. Электронегативные мозговые волны оценивались, усреднялись и интегрировались с помощью компьютера системы LINC–8. |
Шейбел и Шейбел (1967, Рис. XV–3 и XV–4) также описали эту систему нейронов; по мнению этих авторов, она функционирует как система неспецифических афферентных путей к коре больших полушарий. Характерно, что аксоны этих клеток длинные и на их окончаниях имеются разветвления, обвивающие апикальные дендриты пирамидных клеток, свободно соединяющиеся с дендритами, в противоположность хорошо ограниченным синапсам, типичным для специфических афферентов. Эти данные, а также эксперименты Фукса, Гамбургера и Хокфельта (1968), показавшие, как отмечает Стайн, что неспецифические соединения являются в большинстве случаев адренэргетическими (то есть, что аксоны и их клетки содержат норадреналин), говорят о том, что эта система нейронов ответственна за запуск индуктивных процессов посредством стимуляции РНК и облегчения синтеза протеина. Стимулирующее влияние норадреналина на синтез, протеина, по-видимому, опосредствовано циклом адениномонофосфата (см. «The Role of Cyclic AMP in the Nervous System» in «Neurosciences Research Program Bulletin», July 1970, Vol. 8). Кити как-то отметил: «Интересно, что стимуляция протеинокиназы посредством адениномонофосфата может значительно усиливаться с помощью ионов магнезии или калия и тормозиться кальцием, что показывает, каким образом эффект адренэргической стимуляции может по-разному воздействовать на умеренно активные и неактивные синапсы!» Я подчеркнул бы ещё и особое значение нейронной матрицы, на которой реализуется аминэргический подкрепляющий эффект. Эта матрица (нейронная сеть), чувствительная главным образом к холинэргическим веществам, и создаёт то, что в главе XIV описывалось как компетентность мозговой ткани. Существуют данные, говорящие (см. Pис. XV–5) о том, что адренэргическая стимуляция оказывает влияние на холинэргический механизм или по крайней мере — на активирующие и тормозящие механизмы мозгового ствола. Вспомним также, что в экспериментах Креча, Розенцвейга и Беннета (см. Pис. П–6) было обнаружено, что ацетилхолин участвует в эффекте влияния обогащённой среды на рост коры. Могут ли запускаться эти более продолжительные холинэргические процессы с помощью адренэргического механизма подкрепления или эти процессы протекают независимо от последнего? Гликман и Шифф (1967) только на основе обширного обзора результатов, наблюдаемых в экспериментах по изучению влияния электрической стимуляции мозга на поведение, дали частичный ответ на этот вопрос. В этом обзоре выделяется два типа врождённых ответов, один из них состоит в «приближении», а другой в «удалении» от объектов-стимулов. Авторы установили корреляцию между участком электрической стимуляции в стволе мозга, откуда запускается подобный тип поведения, и положительным и отрицательным подкрепляющими эффектами при электрическом самораздражении (см. Pис. V–1 и V–2). Они также высказали предположение относительно некоторых механизмов, посредством которых контролирующие кортикальные влияния (тормозящие и облегчающие) могут регулировать работу центрального эффекторного механизма ствола мозга. Однако они не поняли важности проблемы модификации поведения посредством подобных контролирующих корковых влияний. Это объясняется тем, что авторы не располагали фактами, которые указывали бы на вариабельную природу механизмов, приводящих к реакции (см. Главу XIV), в связи с чем возникает необходимость в особом, способном к модификациям состоянии, имеющем интегративные свойства. В настоящее время анатомическая основа подобных способных к модификации интегративных процессов найдена благодаря открытию системы адренэргических афферентов, берущих своё начало в стволе мозга и достигающих апимальных дендритов. коры больших полушарий. Факты, на которых основываются гипотезы о сущности процесса индукции, получены почти исключительно в опытах на животных. Однако недавний обзор проблемы мозговых поражений у человека, вызывающих серьёзные нарушения процессов памяти, неожиданно подтвердил существование механизма, подобного тому, который здесь описан. Ограниченные поражения коры больших полушарий (включая гиппокамп) или базальных ганглиев не всегда коррелируют с нарушениями памяти. Однако г когда патологический процесс затрагивает структуры ствола мозга, у больных наблюдается «неспособность к образованию новых следов памяти, то есть антероградная амнезия (когда продолжительное заучивание не помогает)», несмотря на «сохранность активного состояния внимания, уровня бодрствования-мозга» и «нормальную способность мыслить, решать задачи и так далее»; в таких случаях при выздоровлении интеллектуальная деятельность лишь «немного ухудшается по сравнению с нремор-бидным уровнем, несмотря на грубые нарушения памяти». Анатомически «ядра, разрушенные поражением, во всех случаях располагаются по средней линии, а именно в 2–3 мм от эпендимы. Иными словами, их расположение в зоне третьего желудочка (и сильвиева водопровода) оказывается более важным как фактор, определяющий их чувствительность, чем все другие свойства этих ядер» (Adams, 1969. — p. 98–102). Нейрофизиологи также внесли известный вклад в изучение мозговых механизмов, определяющих процессы индукции как у человека, так и у животных. Несколько лет назад Грей Уолтер (Walter et al., 1964; Walter, 1967) проанализировал медленно протекающие изменения электрической активности мозга в различных ситуациях. Он обнаружил отрицательное отклонение электрического потенциала при действии одного сигнала, подготавливающего организм к Ответу на следующий, ожидаемый стимул. В наших экспериментах (Donchin et al., 1971; Рис. XV–6 и XV–7) подобные отрицательные отклонения вызывались с помощью разнообразных подготовительных состояний ожидания и место возникновения изменений потенциала зависело от типа задания, требующего этого подготовительного состояния, например: если обезьяна или человек готовится, ничего не делая, по выполнению ответа, максимальная CNV (контингентная негативная вариация) появляется в лобной коре; если требуется подготовиться к моторному ответу, отрицательный потенциал появляется прежде всего в моторной коре и предшествует осуществлению движения; если требуется выполнить длительное ответное действие, например продолжительный подготовительный нажим на рычаг, максимальный отрицательный потенциал возникает в соматосенсорной коре. Когда же наконец появляется ожидаемый второй стимул, отрицательный потенциал завершается резким положительным отклонением. В других экспериментах в ситуациях выработки оперантных условных рефлексов эти завершающие позитивные электрические потенциалы были связаны с появлением подкрепляющего агента (Clemente et al., 1964; Grandstaff, 1969; Lindsley, 1969). Таким образом, CNV с её завершающим быстрым положительным потенциалом сигнализирует об ожидании его завершении, о подготовительных фоновых процессах, определяющих судьбу стимула, которые обсуждались в главе XIV. Непосредственный эффект может быть получен также с помощью отрицательной (катодной) и положительной (анодной) электрической стимуляции коры больших полушарий во время научения (Stamm, 1961; Morrell, 1961). Отрицательная стимуляция повышает скорость научения, положительная оказывает противоположное влияние. Все эти эксперименты дают убедительное доказательство того, что изменения электрического потенциала возникают в мозгу и до и во время подкрепления. Однако мы фактически ничего не знаем о тех механизмах, которые вызывают эти изменения электрического потенциала. Столь же мало знаем мы и о нейрохимических процессах, сопровождающих эти электрические изменения. Возможно, что CNV отражает или мобилизует «холинэргическую компетентность», а положительные потенциалы возникаюг вследствие активации адренэргических путей или вызывает эту активацию. Тот факт, что электрические изменения связаны с подкреплением, открывает пути для изучения данной проблемы на поведенческом, гистологическом и химическом уровнях — пути, которые в течение ряда лет были полностью закрыты. Понимание подкрепления как процесса индукции выдвигает гипотезы, которые могут быть проверены в биологических исследованиях и помогут разгадать ту загадку, над которой полвека безуспешно бились бихевиористы. Регистрация подкрепления |  | | Рис. XV–8. Среднее числа кожногальванических реакций, возникающих за 10-секундный период между включением света и его выключением в первых 40 и во вторых 40 пробах по каждой группе обезьян. Нормальные контрольные животные научаются предвосхищать выключение света, а у животных с удалённым миндалевидным ядром эта способность нарушается (Bagshaw and Coppock, 1968). |  | | Таблица XV–2. Влияние удаления ампгдалы на перенос навыка иа новое но связанное с прежним задание» (см. объяснение к Рис. XI–3). Обезьяны после удаления ампгдалы обучаются, выполнению второго задания, как совершенно нового, тогда как нормальные контрольные животные действуют в этих условиях на основе своего прошлого опыта (Schwartzbanm, Pribram, 1960). |
Вспомним ещё раз эксперименты с угашением (см. Главу III, XI), Регистрируя ряд физиологических индикаторов, мы видим, что — организм отвечает на новый стимул ориентировочной реакцией. Эта реакция угасает при повторном предъявлении стимула. Вспомним также, что после удаления миндалевидного ядра физиологические индикаторы ориентировочного рефлекса, такие, как кожногальваническая реакция (КГР), изменение частоты сердцебиений, дыхания, не обнаруживаются, несмотря на то, что — в других условиях эти физиологические ответы остаются сохранными. Как это ни парадоксально, поведенческое выражение ориентировочной реакции в этих случаях не только сохраняется, но и не угасает. Я интерпретирую этот факт таким образом, что после амигдалэктомии ориентировочный процесс не регистрируется в мозгу так, как это происходит в норме. В ситуации выработки условных рефлексов также отсутствует регистрация этих показателей, как это было обнаружено в опытах с обезьянами, перенесшими подобную операцию (Bagshow and Coppock, 1968; Рис. XV–8). Если у нормальных животных наблюдается постепенное увеличение числа электрофизиологических ответов, возникающих во время предъявления стимула, и удлинение периода, во время которого появляются предвосхищающие ответы, то у оперированных животных подобных предвосхищающих КГР и увеличения их числа не наблюдается. Эти результаты говорят о том, что миндалевидное ядро интимным образом связано с организацией процессов во времени, обеспечивая процессы, участвующие в ответе на повторные события, иначе говоря, процессы серийной регистрации в памяти значимых событий. В нормальном организме существует как бы «внутреннее эхо» событий, без которого соответствующая регистрация не происходит. То, что миндалевидное ядро участвует в процессе усвоения опыта, подтверждает и поведение животных (Douglas and Pribram, 1966). В простых задачах на различение часто повторяющихся объектов эти затруднения в регистрации событий почти не заметны. Однако, если порядок подкрепления изменяется таким образом (Barrett, 1969), что начинает подкрепляться ранее не подкреплявшийся стимул (переделка дифференцировки) или вводится новый стимул, как в том эксперименте, когда обезьяна должна была использовать опыт, полученный в другой, сходной, но не идентичной ситуации, животные, перенесшие амигдалэктомию, не могут решить задачу (Schwartzbaum and Pribram, 1960; Рис. XV–9; Bagshow, Pribram, 1965). В таких опытах обезьяны, подвергшиеся амигдалэктомии, обнаруживают повышенную реактивность на новизну (по сравнению с контрольными животными) и эта реактивность сохраняется длительное время после того, как нормальные животные уже привыкли к ситуации. 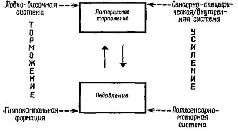 | | Рис. XV–9. Модель коркового контроля над афферентными подавляющими и тормозными процессами (в лобно-височную систему включается и миндалевидное ядро). |
Эти результаты, а также результаты, полученные при изучении физиологических индикаторов ориентировочного рефлекса, говорят о том, что миндалевидное ядро обычно контролирует «контрастное усиление» нервных процессов, нервные механизмы латерального торможения во входных каналах, принимающих информацию (что происходит всегда, если работает много нейронных аппаратов). Действие той системы, частью которой является амигдалярный комплекс, состоит в подавлении эффекта разбалансировки, возникающего при быстром изменении ситуации. Эти изменения ситуации приводят к смене режима работы: самоподавление возбуждения сменяется латеральным торможением в том широко распространённом в нервной системе реципрокном содружестве, которое составляет нейронный тормозный механизм — своего рода «заслонку» в системах «входа» (см. Pис. XV–9)… Короче говоря, наличие нормально функционирующего миндалевидного ядра, по-видимому, вводит в действие «внутреннее эхо», что и приводит к регистрации соответствующих, то есть совпадающих с контекстом, событий; это происходит с помощью подавления эффекта воздействия каждого нового стимула, вызывающего значительную разбалансировку организма. Сам факт регистрации событий посредством своего рода «внутреннего эха», удлиняющего время воздействия на мозг совпадающих событий, наводит на мысль о нейронном механизме индукции. Работу амигдалярной системы можно понимать как усиление процесса индукции посредством предохраняющей интерференции (Pribram, Douglas and Pribram, 1969) и, таким образом, предоставление достаточного времени для того, чтобы «внутреннее эхо» могло проявиться. Только тогда, когда эти процессы интерференции контролируются, и может осуществляться завершение поведенческого акта и поведение непрерывно регулируется. Управление правильными и ошибочными ответамиПомимо механизма регистрации, существует ещё один наиболее важный механизм, обеспечивающий длительное управление поведением, который мы называем завершением действия (commitment). Этот более тонкий процесс относится к тем явлениям, которые математические психологи называют «негативными случаями». Этот процесс следует за самой ранней фазой процесса регистрации и характеризуется началом активного игнорирования организмом тех явлений, которые уже происходили однажды, во теперь больше не подкрепляются. Имеются надёжные доказательства того, что в этом процессе участвует гиппокамп, часть переднего мозга, примыкающая к миндалевидному ядру. 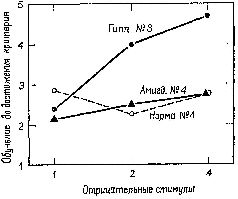 | | Рис. XV–10. График результатов экспериментов с изменением количества отрицательных стимулов в задачах на различение (Douglas et al., 1969). |
Так, например, мы показали, что, если в ситуации различения число неподкрепляемых (то есть негативных) стимулов варьирует от одного до четырёх, выполнение задания нормальными животными почти не изменяется. Однако после удаления гиппокампа обезьяны значительно дольше обучаются заданию, в котором предъявляются четыре неподкрепляемых стимула, и скорость их научения пропорциональна числу подобных неподкрепляемых агентов (Douglas et al., 1969; Рис. XV–10). Затем нормальные животные переходят ко второй фазе обучения: они учатся игнорировать новые неподкрепляемые стимулы. У обезьян, перенесших операцию удаления гиппокампа, механизм этой второй фазы нарушается; они не способны оценивать свои ошибки, если таковые делаются вследствие неподкрепления. Эта неспособность к оценке ошибок особенно заметно проявляется в тех ситуациях, когда требуется подавить предварительно заученные формы поведения (Douglas, 1967, Kimble, 1969). Однако изучение такого подавления выдвигает свои проблемы, которые составляют содержание следующей главы. 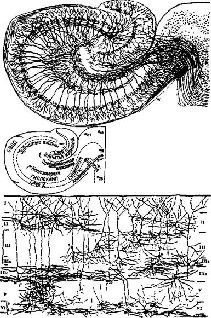 | | Рис. XV–11. Цитоархитектоника гиппокампальной извилины: зубчатая извилина и гиппокамп. Верхний рисунок из работы Кахала (1911), нижний — из работы Лоренте де Но (1949). |
Механизм, обеспечивающий активное игнорирование ошибок, несомненно, действует в более широком диапазоне ситуаций, чем те, которые изучаются в лабораторных условиях. Во всех подобных ситуациях должна совершаться быстрая оценка соответствия наличных стимулов прежним результатам поведения (прежним подкреплениям). Структура коры гиппокампа, столь сходная во многих отношениях со структурой мозжечка, анатомически очень подходит для этой цели (см. Pис. XV–11). Мгновенный подсчёт вероятности ошибки или рассогласования (основанный на оценке текущих состояний гомеостазиса и влияния на него результатов выполнения последней или нескольких последних задач) может, следовательно, служить в качестве контекста (фона), по отношению к которому и определяется это соответствие, и таким образом может быть оценена правильность ответа. С этой точки зрения гиппокамп можно рассматривать как механизм регуляции подкрепления и выполнения задачи, аналогичный мозжечку. Теперь обратимся к третьей группе проблем, связанных с модификацией поведения: модификации в процессе выполнения задания. В главе XIV рассматривалась компетентность организма в процессе научения. В настоящей главе мы уточнили нервные механизмы процессов подкрепления, обеспечивающие завершение действия, и рассмотрели мозговые механизмы, участвующие в завершении задачи. Однако завершение задачи ещё не означает достижение результата. В следующей главе мы покажем, как завершённые действия превращаются в акт поведения. РезюмеДлительные изменения компетентности могут возникать благодаря процессу, сходному с тем, который стимулирует эмбриональные ткани к дифференциации. Характерные черты эмбриональной индукции и поведенческих процессов, связанных с подкреплением, удивительно сходны. Факты показывают, что адренэргические химические процессы индукции стимулируют образование РНК и синтез протеина в механизме соединений весьма разветвлённой нервной системы. Механизм индукции нуждается в защите от вмешательства на всех стадиях развития, иначе не будет получен нужный эффект. Большое число мозговых процессов обеспечивает завершение серии непрерывно подкрепляемых звеньев поведенческого акта после того, как они уже введены в действие. |